Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Dabing Zhang and Version 2 by Camila Xu.
Sucrose is produced in leaf mesophyll cells via photosynthesis and exported to non-photosynthetic sink tissues through the phloem. The molecular basis of source-to-sink long-distance transport in cereal crop plants is of importance due to its direct influence on grain yield—pollen grains, essential for male fertility, are filled with sugary starch, and rely on long-distance sugar transport from source leaves. Here, we overview sugar partitioning via phloem transport in rice, especially where relevant for male reproductive development.
- sugar partitioning
- phloem
- sink
- source
- sugar signaling
1. Introduction
Rice (Oryza sativa), the monocot model plant, is a major crop, meeting the food demands of more than 50% of the global population [1][2][1,2]. Reproductive development, which connects the dominant diploid sporophytic and short haploid gametophytic stages, is a critical element in grain production [3]. The male reproductive organ, the stamen, consists of a filament and an anther containing multiple specialized tissues that generate mature male gametophytes, the pollen grains, via a series of developmental events such as meristem specification, cell differentiation, meiosis, mitosis, and starch accumulation [4][5][4,5].
Sugars are the constituents of main anther, and play essential roles in cell structure formation, energy supply, and male fertility in response to environmental conditions [6][9]. In rice, the expression of Cell Wall Invertase 3 (OsCWIN3/OsINV4) correlates with sucrose accumulation and pollen sterility depending on temperature [7][10], while two MYB domain proteins, Carbon Starved Anther (CSA) and CSA2, regulate sugar partitioning and male fertility in response to photoperiod [8][9][10][11][11,12,13,14]. The sugar transporter OsXa13/OsSWEET11 plays essential roles in pollen development and disease resistance against bacterial blight [12][13][15,16].
2. Strategies of Source-to-Sink Sugar Partitioning
Carbon is fixed from carbon dioxide into carbohydrate in chloroplasts of leaf tissues, primarily mesophyll cells, and accumulated in the cytosol of the same cells. The energy demands of sink tissues, such as roots, flowers, and seeds, drive the export of sugars from the leaf, mainly in the form of sucrose, via long-distance transport in plant vasculature, the phloem [14][17]. Over half of the photo-assimilates (50–80%) are exported from source leaves to maintain non-photosynthetic sink tissues [15][18]. Carbohydrate partitioning from source-to-sink tissues comprises three elements [16][19]: phloem loading of sugars from source tissues; transportation in the sieve element of the phloem; phloem unloading of sugars to sink tissues [17][20]. Phloem is composed of several cell types, including parenchyma cells, sieve elements (SEs), and companion cells (CCs) [18][19][21,22]. Phloem loading is the first vital step in sugar’s long-distance transport—transferring the sugars from mesophyll cells to the SEs and CCs of the phloem [20][21][22][23,24,25]. Three different strategies are used for phloem loading by different plants according to the abundance of plasmodesmata, SUT activity, and the concentration gradient of photosynthates (Figure 1) [23][26].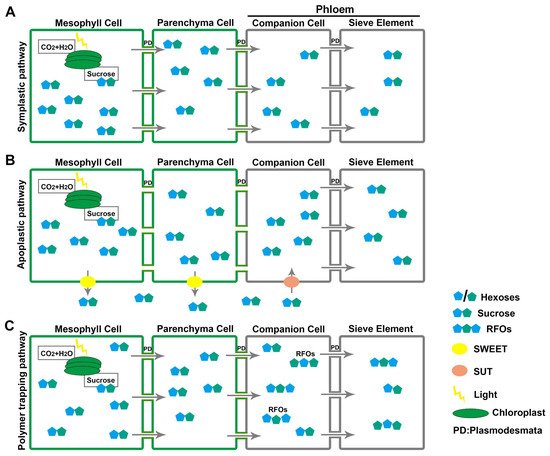
Figure 1. Three strategies for phloem loading. (A) Symplastic pathway: sucrose accumulates in mesophyll cells and is passively translocated to the phloem through plasmodesmata (PD) along the concentration gradient. (B) Apoplastic pathway: sucrose is exported to the apoplast by SWEETs and, after diffusion, imported into the phloem by SUTs. (C) Polymer trapping: sucrose is passively exported to phloem companion cells and synthesized into RFOs that can only move into sieve element cells due to their larger molecular mass.
3. Proteins Involved in Sugar Partitioning
3.1. Sucrose Transporters (SUTs)
Sucrose transporters (SUTs) act as symporters to import the sucrose from the apoplasm into phloem CCs against the concentration gradient, driven by the motive force generated by H+-ATPases (Figure 1B) [25][27][28,32]. The 12 transmembrane domains of the SUT protein forms a pore to transport sucrose across the plasma membrane [28][33]. The first sucrose transporter (SoSUT) was found in spinach (Spinacea oleracea) by an elegant yeast complementation strategy [29][34]. Nine and five SUTs have been found in Arabidopsis and rice, respectively [30][31][35,36]. Based on sequence, sub-cellular location, and activity, SUTs have been classified into three types: type I (specific to eudicots, plasma membrane–localized); type II (present in all plants, plasma membrane–localized); and type III (present in all plants, vacuolar membrane–localized) [32][37]. In rice, OsSUT1, OsSUT3, OsSUT4, and OsSUT5 are type II SUTs, and OsSUT2 is a type III tonoplast SUT (Table 1) [32][37].Table 1.
Proteins involved in sugar metabolism in rice.
| Gene Family | Number of Genes | Reported Genes/Reference |
|---|---|---|
| SUT | 5 | SUT1 [33][34][38,39]; SUT2 [35][40]; SUT3 [36][41]; SUT4, SUT5 [30][35] |
| SWEET | 21 | SWEET5 [37][42]; SWEET6a [38][39][43,44]; SWEET6b [39][44]; SWEET11 [12][15]; SWEET14 [13][16]; SWEET15 [40][45] |
| MST | 64 | |
| AZT subfamily | 6 | AZT3 [10][13] |
| ERD subfamily | 6 | |
| pGlcT subfamily | 4 | |
| Xylose subfamily | 2 | |
| STP subfamily | 15 | MST1, MST2, MST3 [41][46]; MST5 [42][47]; MST6 [43][48]; MST8 [8][11]; SPT17, SPT27 [38][39][43,44] |
| PLT subfamily | 28 | |
| INT subfamily | 3 | |
| Invertases | 18 | |
| VIN | 2 | VIN2 [44][49] |
| CIN | 8 | CIN8/Cyt-INV1 [45][50] |
| CWIN | 8 | GIF1 [46][51]; INV4 [7][10]; CWIN6 [39][44] |
The shaded areas represent subfamilies of these proteins.
3.2. Sugars Will Eventually Be Exported Transporters (SWEETs)
SWEETs are a group of evolutionally conserved genes expressed in eukaryotes, prokaryotes, and archaea [47][48][52,53]. These genes, encoding MtN3/saliva domain proteins, were initially found to encode glucose transporters [49][54], and have since been found to be capable of transporting a variety of mono- and di-saccharides [37][50][51][52][42,55,56,57]. According to their protein structures, SWEET proteins encode either one or two MtN3/saliva domains [53][58]. Rice encodes 21 SWEET proteins that are involved in multiple biological processes (Table 1) [53][58]. OsSWEET11, containing two MtN3/saliva domains, acts as a glucose uniporter in panicles and anthers [12][15]. Its knockdown mutant reveals defects in microspore development, suggesting a function in male development [12][15]. OsSWEET11 is also upregulated in response to bacterial infection by Xanthomonas oryzae pv. oryzae [12][15]. OsSWEET14 has a similar disease response, and its knockout mutant showed growth retardation, reduced plant size, and insensitivity to bacterial infection [13][16].3.3. Invertases (INVs)
Invertases (INVs) encode proteins that hydrolyze sucrose into glucose and fructose [54][60], classified according to sub-cellular location into vacuolar (VIN), cell wall (CWIN), or cytoplasmic (CIN) invertases (Table 1) [55][56][61,62]. CINs prefer a neutral pH of 7.0–7.8 in the cytosol, while VINs and CWINs have an optimal pH of 4.5–5.5 [55][61]. Rice has 19 invertase genes, including nine CWINs, two VINs, and eight VINs (Table 1) [57][63]. CWIN proteins bind to the cell wall and play essential roles in sugar transmembrane transport during phloem unloading [55][61].3.4. Monosaccharide Transporters (MSTs)
Monosaccharide transporters (MSTs) are membrane proteins involved in the transmembrane transport of hexoses, hydrolyzed from sucrose by INVs, in sink tissues in the apoplastic pathway (Figure 1B) [58][64]. An Arabidopsis phylogeny of 53 MST proteins suggests seven subfamilies—AZT, XTPH, ERD, pGlcT, PLT, INT, and STP (Table 1)—many of whose expression patterns or function have not yet been characterized [59][65]. Among the seven subfamilies of MST proteins, AZT and XTPH proteins localize on the tonoplast and play essential roles in sugar transport to the tonoplast [60][61][62][66,67,68]. AtERD6, a member of ERD proteins, was proved to be involved in the transport of monosaccharides, whose expression was induced by abiotic stress [63][69]. pGlcT proteins are transporters of glucose, and PLT proteins are symporters of polyols and monosaccharides [64][65][70,71]. AtINT4, the first identified member of the INT proteins, exhibits H+ symporter activities for myoinositol in yeast (Saccharomyces cerevisiae) and Xenopus laevis oocytes [66][72]. MST members of the STP sub-family are H+/hexose cotransporters locating on plasma membranes, which transport a series of hexoses, including glucose, fructose, galactose, xylose, mannose, pentose, and ribose [58][64].4. Roles of Sugar Transporters in Phloem Loading and Unloading
Photosynthesis—“source activity”—and sink energy utilization—“sink strength”—combine to raise plant productivity [17][67][20,73]. Understanding the processes of phloem loading in source leaves and unloading in sink tissues can improve source activity and sink strength, leading to higher grain yields (Figure 2). After long-distance phloem transport from source tissues, sugar (mainly sucrose) is unloaded in sink organs; however, this process will lead to sucrose accumulation in sink tissues (reduced sink strength), resulting in reduced efficiency in sugar transport and source activity (Figure 2) [68][74].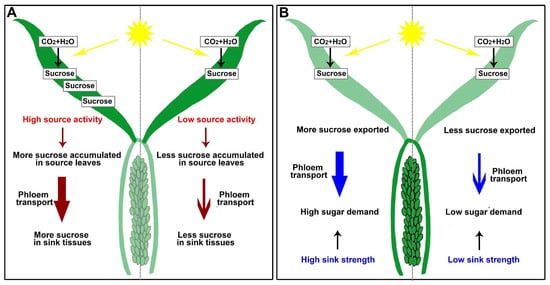
Figure 2. Schematic diagrams of sugar source-to-sink transport in rice. (A) High source activity in source leaves promotes phloem transport. (B) High sink strength results in high sugar demand, increasing the sugar transport.