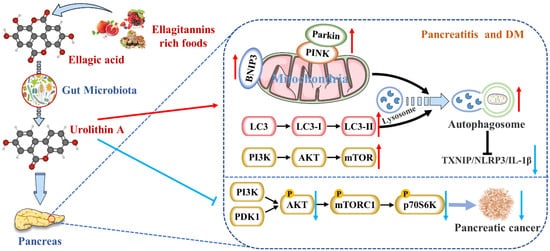
Urolithin A (Uro A) is a dietary metabolite of the intestinal microbiota following the ingestion of plant-based food ingredients ellagitannins and ellagic acid in mammals. Accumulating studies have reported its multiple potential health benefits in a broad range of diseases, including cardiovascular disease, cancer, cognitive impairment, and diabetes.
1. Reduces the Expression of Pancreatic Inflammatory Factors
The inflammatory microenvironment of the pancreas led to pancreatitis and was the main reason for the decline in endocrine function
[1][ 83 ]. Some researchers suggested that if
β cells would express high levels of NF-κB signaling marking, cells’ proliferative and regenerative potential were reduced. The NF-κB-expressed β cells also emerged with a premature upregulation of
socs2, a gene that inhibits proliferation
[ [2].84 ]。It has been widely reported that EA can inhibit pancreatic inflammation (
Table 1). In an experimental model of spontaneous chronic pancreatitis, male Wistar Bonn/Kobori rats were fed a diet supplemented with EA (100 mg/kg body weight/day) for ten weeks. They found that EA attenuated pancreatic inflammation and fibrosis by increasing pancreatic weight and decreasing MPO activity (a neutrophil infiltration index), collagen content, transforming growth factor-β1 (TGF-β1) expression, activated pancreatic stellate cells (PSCs), and ED-1-positive cells
[3].[ 85 ]。Masamune et al. also reported that EA inhibited the production of monocyte chemoattractant protein-1 (MCP-1) and activation of activator protein-1 (AP-1) and MAPK in PSCs, all induced by interleukin (IL)-1β and TNF-α
[4].[ 86 ]。Meanwhile, EA inhibited PDGF-BB-induced tyrosine phosphorylation of PDGF P-receptors and the downstream ERK and Akt activation in PSCs. In particular, EA inhibited ROS production in PSCs in response to TGF-β1 or platelet-derived growth factor (PDGF)
[ [5].87 ]。
Table 1. Potential effects and mechanisms of urolithin A (Uro A) and its precursor compounds EA on pancreatic diseases.
| Disease Model |
Treatment |
Metabolic Response |
Ref. |
| Spontaneous CP in male Wistar Bonn/Kobori rats |
100 mg/kg BW/day orally administered with EA for 10 weeks |
-
Attenuated pancreatic inflammation and fibrosis
-
Increased pancreatic weight
-
Decreased MPO activity, collagen content, TGF-β1 expression; activated PSCs and ED-1-positive cells
|
[3][ 85 ] |
| PSCs were isolated from rat pancreas tissue to culture activated, myofibroblast-like phenotype |
1–25 μg/mL EA |
-
Inhibited PSCs’ proliferation and migration
-
Inhibited MCP-1 production and the expression of smooth muscle actin and collagen genes
-
Inhibited the tyrosine phosphorylation of PDGF P-receptor
-
Inhibited the activation of Akt and MAPKs
|
[4][ 86 ] |
| L-arginine induced AP in rats |
85 mg/kg orally administered with EA |
-
Decreased TOS levels
-
Increased TAC levels
-
Decreased TNF-α, IL-1β, and IL-6 serum levels
-
Healed inflammatory and oxidative stress
|
[5][ 87 ] |
| MIN6 β-cell inflammations were induced using 25 mM glucose and 0.5 mM palmitic acid |
Uro A |
|
[6][ 88 ] |
| Alcohol-associated CP in C567 BL6/J mice |
Administered during the last 3 weeks of alcohol-associated CP induction |
|
[7][ 89 ] |
| DM in male C57 BL/6 mice was achieved by a HFD and intraperitoneal STZ injections |
50 mg/kg BW/day orally administered with Uro A for 8 weeks |
|
[8][ 90 ] |
| Human PDAC cell lines; PDAC mice were achieved by injecting PANC1 cells into the flank of 6-week-old Fox1-nu/nu mice |
0–100 μM; 20 mg/kg BW/day (5 days/week) orally administered with Uro A |
-
Inhibited the proliferation and migration of PDAC cells
-
Enhanced apoptosis by down-regulating the PI3K/AKT/mTOR pathway
-
Inhibited the PDK1/AKT/mTOR pathway
-
Reduced immunosuppressive TAMs and regulatory T cells
|
[9][ 91 ] |
| PKT (Ptf1acre/+; LSL-KrasG12D; Tgfbr2fl/fl) mice, an aggressive genetically engineered PDAC mouse model |
Orally administered with Uro A for 5 weeks |
-
Inhibited AKT, PS6K, and STAT3 signaling
-
Reduced the Ki67-positive tumor cells
-
Increased cleaved caspase-3 expression
|
[10][ 92 ] |
| Neonatal STZ-induced non-obese T2DM rats |
25–100 mg/kg BW orally administered with EA |
|
[11][ 93 ] |
Although EA had many promising developments, it was poorly absorbed in the human gut, limiting its anti-inflammatory effects. As mentioned above, EA was metabolized by microorganisms into a series of downstream compounds, such as Uro A
[ [12]5 ]. A well-known effect of preclinical models exposed to Uro A was the attenuation of harmful inflammatory responses
[13][ 94 ]. Uro A showed more potent anti-inflammatory properties than EA or ETs, suggesting that it might be the main compound for treating AP or CP (
Table 1). The anti-inflammatory effects were first reported to reduce the mRNA and protein levels of inflammatory marker COX-2 in rats with acute colitis
[ [14]95 ]. Zhang et al. had firstly reported that Uro A inhibited the thioredoxin-interacting protein (TXNIP)/Nod-like receptor family pyrin domain containing 3 (NLRP3)/IL-1β inflammation signal in MIN6 β cells by modulating AMPK (
Figure 12)
[6][ 88 ]. Finally, they testified that Uro A also down-regulated the protein kinase RNA (PKR)-like ER kinase (PERK) and promoted AMPK phosphorylation
[15][ 96 ]. The latest research showed that Uro A can attenuate the severity of alcohol-associated chronic pancreatitis (ACP) in C56BL6/J mice by regulating the PI3K/AKT/mTOR signaling axis
[7][ 89 ].
Figure 12. Uro A is metabolized by gut microbiota after ingestion of ETs and EA in mammals and has multiple potential health benefits. Uro A can attenuate pancreatic diseases by inhibiting inflammatory signaling pathways, activating autophagy, maintaining the mitochondrial function, and improving the immune microenvironment.
Nevertheless, studies on reducing pancreatic inflammation by Uro A had only been verified in animals and cells without clinical studies. The upstream mediators of Uro A’s anti-inflammatory effects, including the NF-κB and AhR-Nrf2 pathways, were mainly studied in vitro [16][36]. Nevertheless, the mechanisms of Uro A action in the context of inflammation seemed to vary with tissues and conditions. Hence, the differences in Uro A’s mitigation degree and mechanism on AP and CP need to be further explored.
2. Activates Autophagy and Maintains Mitochondrial Function in the Pancreas
Mitochondrial damage, such as the loss of mitochondrial DNA (mt DNA) integrity, the alteration of mitochondrial morphology, and dysfunction, can lead to cellular senescence and apoptosis
[17][ 97 ]. On the one hand, mitochondria acted as both nutrient sensors and signal generators for insulin secretion in β cells. Moreover, nutrients can inhibit the ATP-sensitive K
+ (K
ATP) channel and then enhance insulin secretion either by acting as substrates for mitochondrial ATP synthesis (the triggering pathway) or by regulating Ca
2+ channels (the amplifying pathway). On the other hand, mitochondria were the primary source of reactive oxygen species (ROS) at the level of the electron transport chain so that mitochondria might be the main targets of ROS damage
[ [18]98 ]. Additionally, many studies have revealed a causal relationship between pancreatic diseases and dysregulation of mitochondrial dynamics (including fusion and fission)
[17][19][20][ 97 , 99 , 100 ]. Thus, mitochondrial damage gave rise to decreased pancreatic function. The most consistent effect of Uro A across species, including cells, worms, mice, and humans, was improved mitochondrial health
[13][ 94 ]. This benefit was driven by the clearance and recycling of dysfunctional mitochondria, known as selective autophagy
[ 101 [21]]. For example, Uro A increased the expression of mitochondrial autophagy genes
lgg-1 lgg-1,
pink-1, pink-1, and
pdr-1 pdr-1, encoding for LC-3B, and formation of autophagosome vesicles in C. elegans
[ [22]29 ].
Interestingly, Pink1 knockdown in microglia eliminated Uro A-mediated reductions in TNF-α and increased IL-10, suggesting that Uro A reduces neuroinflammation by inducing mitochondrial autophagy
[23][ 34 ]. Zhang et al. also proved that Uro A inhibited glucolipotoxicity-induced ER stress and the TXNIP/NLRP3/IL-1β inflammation signal in MIN6 β cells by modulating autophagy
[6][ 88 ]. Remarkably, the inhibitory effects of Uro A on p62 were stronger than TXNIP-inhibitor verapamil (
p p <
0.05)
[ [24]102 ]. Uro A promoting PINK1/Parkin-mediated mitophagy was also reported in pancreatic cells of diabetic mice
[ [8]90 ]. Therefore, Uro A restoring the correct level of mitochondrial autophagy to maintain normal mitochondrial function is highly likely to be the mechanism of Uro A reducing pancreatic diseases (
Figure 12).
3. Inhibits Endoplasmic Reticulum Stress in the Pancreas
The misfolding and inhibition of protein folding in the endoplasmic reticulum (ER) lead to the aggregation of unfolded proteins, resulting in ER stress
[25][ 103 ]. Li et al. showed that the ER stress and unfolded protein response (UPR) accompanied by the accumulation of protein aggregates emerged as a significant pathway affected by aging, specifically in β cells. Simultaneously, the transcriptomic dysregulation of UPR components was linked to activating transcription factor 6 (ATF6) and inositol-requiring enzyme 1 (IRE1) signaling pathways
[26][ 8 ]. ER stress-related apoptosis lead to a reduction in β-cell proliferation and regeneration, ultimately resulting in reduced insulin secretion and increased T2DM morbidity
[27][ 104 ]. Therefore, maintaining transcriptional stability and reducing protein homeostasis loss during aging was crucial to recovering pancreatic function. It has been reported that Uro A suppresses glucolipotoxicity-induced ER stress in pancreatic beta cells
[6].[ However,88 more studies are needed on Uro A’s upstream and downstream pathways in the pancreas to improve ER stress.]。However, more studies are needed on Uro A’s upstream and downstream pathways in the pancreas to improve ER stress.4. Inhibits the Occurrence and Development of Pancreatic Tumors
High intakes of berries rich in ETs, including strawberries, pomegranates, and blueberries, were inversely associated with PDAC incidence
[28][ 105 ]. EA, an intestinal metabolite of ellagic tannins, inhibited multiple carcinogenic pathways activated in PDAC, such as COX-2, NF-κB, and Wnt signaling, so that EA successfully arrested cell cycles and reversed epithelial to mesenchymal transition in PDAC
[29][ 106 ]. As a downstream compound of EA, Uro A showed more potent antioxidant and anti-inflammatory properties, improving bioavailability and anti-tumor effect
[30][ 107 ]. It has been demonstrated that the S473 phosphorylation site of AKT is activated by PI3K
[31][ 108 ]. Uro A treatment resulted in a dose-dependent reduction in phospho-AKT (p-AKT) expression in PDAC cell lines, leading to a significant down-regulation of phospho-p70 S6 kinase (p-PS6K) expression regulated by the mTORC1 complex. Therefore, Uro A inhibited the proliferation and migration of PDAC cells and enhanced apoptosis by down-regulating the PI3K/AKT/mTOR pathway
[32][ 109 ] (
Figure 12). Furthermore, Uro A treatment also down-regulated PDK1 (the upstream target of AKT) and p-GSK3β and p-4E-BP1 (the downstream targets of AKT), suggesting that Uro A effectively inhibited the PDK1/AKT/mTOR
[ [33]110 ]. Uro A treatment also reduced immunosuppressive tumor-associated macrophages (TAMs) and regulatory T cells in the engineered PKT mouse model of PDAC. It meant that Uro A treatment attenuated tumor growth and prolonged survival in mice by inducing changes in the immunosuppressive microenvironment of PDAC
[9][ 91 ]. Srinivasan et al. also pointed out that Uro A inhibited AKT, PS6K, and STAT3 signaling, thereby reducing the Ki67-positive tumor cells and increasing cleaved caspase-3 expression in the pancreatic tissues of PDAC mice
[10][ 92 ]. These results suggest that Uro A is a novel inhibitor/regulator for multi-signal pathways in PDAC and has potential in the prevention and treatment of pancreatic cancer (
Table 1).
5. Protects Pancreatic β Cells
There have been many studies on the ameliorative effect of Uro A on DM and its complications. Specifically, Savi et al. first showed that Uro A recovered cardiomyocyte contractility and calcium dynamics in diabetic cardiomyopathy (DCM) rats
[34][ 111 ]. Albasher et al. further demonstrated that Uro A prevents streptozotocin (STZ)-induced DCM in rats by activating SIRT1 expression and deacetylase activity
[35][ 112 ]. Xiao et al. suggested that Uro A can attenuate DM-related cognitive impairment by ameliorating systemic inflammation and intestinal barrier dysfunction through the N-glycan biosynthesis pathway
[36][ 113]. This conclusion was also supported by Lee et al. They pointed out that Uro A prevented DM-associated AD by reducing transglutaminase type 2 (TGM2)-dependent mitochondria-associated ER membrane (MAM) formation and maintaining mitochondrial calcium and ROS homeostasis
[ [37].35 ] Xu et al. indicated that Uro A ameliorated diabetic retinopathy by activating the Nrf2/HO-1 pathway to inhibit inflammation and oxidative stress
[ 114 [38]]. Zhou et al. found that
Phyllanthus emblica Phyllanthus emblica L. facilitated vascular function in STZ-induced hyperglycemia rats by regulating Akt/β-catenin signaling, mediated by the ETs metabolites
[ [39]115 ].
Insulin resistance is one of the core mechanisms of DM. However, as a complex systemic metabolic disease, insulin resistance alone is not enough to cause DM. Islet dysfunction caused by the decrease in the total amount of islet β cells is also the key to the pathogenesis of DM. Studies have shown that β cells in T2DM can be divided into three main states: susceptibility, adaptation, and failure
[40][41][42][ 116、117、118 ]. During insulin resistance, β cells compensate for the dysfunction by increasing insulin demand through insulin secretion
[8][ 90 ]. When β cells fail to compensate for glucose homeostasis, hyperglycemia occurs. More importantly, EA from
Phyllanthus emblica Phyllanthus emblica L. increased the size or number of β cells in diabetic rats. EA also directly increased glucose-stimulated insulin secretion from isolated islets, suggesting that EA acted directly on pancreatic β cells to exert anti-diabetic activity, thereby stimulating insulin secretion and reducing glucose intolerance
[11][ 93 ]. Histopathological results showed that Uro A had protective effects on β cells, such as improving the pancreatic structure and increasing islet size and number. Ultrastructural damages in DM mice pancreas after Uro A treatment, including ER expansion, mitochondria swelling, cristae fracture, and myelin sheath formation, were also significantly improved
[8][ 90].
ThWe
researchers aalso discussed earlier that Uro A prevented β-cell apoptosis in T2DM model mice by activating autophagy and regulating the AKT/mTOR signal
[6][8][24][90,98,99]. However, the specific mechanisms of Uro A improving β-cell structure and function to mitigate DM risk need to be explored further.
In summary, the metabolism and the roles of Uro A in ameliorating pancreatic diseases have been extensively discussed in this section (summarized in In summary, the metabolism and the roles of Uro A in ameliorating pancreatic diseases have been extensively discussed in this section (summarized in
Figure 1). By clarifying Uro A’s metabolism in vivo and Uro A’s mechanisms for protecting the pancreas, it might shed new light on managing pancreatic injuries via plant-based foods rich in ETs and EA.2). By clarifying Uro A’s metabolism in vivo and Uro A’s mechanisms for protecting the pancreas, it might shed new light on managing pancreatic injuries via plant-based foods rich in ETs and EA.