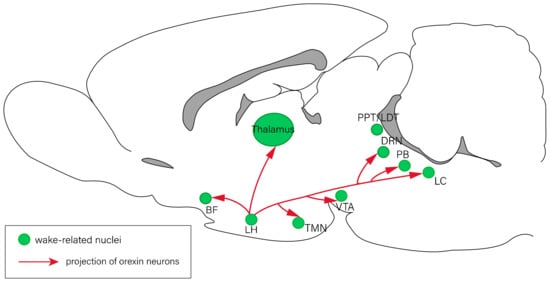
2.2. Orexin Consolidates Wakefulness and Inhibits REM Sleep
Orexin exists in two molecular forms, orexin-A and orexin-B, which both perform physiological functions through the interaction with the GPCRs
[37][41]. Rats have around 3000–4000 orexin-producing neurons in the brain
[38][42], and they are located mainly in the peripherical area of the LH
[39][40][43,44]. These neurons project widely to the CNS, regulating feeding and other behaviors. Many nuclei that regulate sleep–wake behaviors receive projections from the orexin neurons as well, including the LC
[41][45], DRN
[42][46], TMN
[43][47], PPT/LDT
[44][48], and BF
[45][49]. Therefore, orexin is involved in sleep–wake regulation.
2.3. Melanin-Concentrating Hormone Has Positive Influence on Sleep
Melanin-concentrating hormone (MCH) is a cyclic neuropeptide consisting of 19 amino acids
[46][76], which performs physiological functions through the interaction with two GPCRs known as MCH receptor-1 and MCH receptor-2. It serves as an important neuromodulator of homeostasis and performs a large range of integrative functions, which are mainly associated with homeostatic regulation and motivated behaviors
[47][48][77,78]. In mammals, MCH neurons are mainly located in the LH and the zona incerta
[49][50][79,80]. They project to many nuclei that promote REM sleep and arousal, including the LC, DRN, LDT/PPT, and the sub-LDT
[51][81]. Although the location and projection of MCH neurons are remarkably similar to those of orexin neurons
[51][81], they have opposite effects on the modulation of sleep–wake states. MCH neurons have a positive influence on sleep, especially REM sleep
[46][76].
2.4. Neuropeptide S Is Associated with Arousal Induction
Neuropeptide S (NPS) is a peptide composed of 20 amino acids and an endogenous ligand for the NPS receptor. The NPS receptor is a typical GPCR, containing seven membrane-spanning domains. The N-terminal residue of NPS in all species is always serine, hence the name NPS
[52][89]. NPS is expressed in the brainstem, amygdala, hippocampus, and in other regions of the limbic system. Rainer et al. have demonstrated that mRNA expression of the NPS receptor is widespread throughout the CNS, and it is especially abundant in the cortex, thalamus, hypothalamus, and amygdala
[53][90]. In addition, low mRNA levels of the NPS receptor have been observed in the brainstem. In contrast, mRNA expression of the NPS precursor is mainly observed in brainstem nuclei such as the LC and the lateral parabrachial nucleus, while a small number of scattered NPS-positive neurons are found in other brain areas, such as the amygdala and hypothalamus
[52][54][89,91]. NPS induces the mobilization of intracellular Ca
2+ [55][92], increases the intracellular cAMP levels, and stimulates the phosphorylation of mitogen-activated protein kinase
[56][93]. NPS modulates a variety of physiological functions such as food intake
[57][58][59][94,95,96], the regulation of the endocrine system, spatial memory, alcohol seeking, nociception, and anxiety.
2.5. Neuropeptide Y Has a Dual Impact on Sleep–Wake Behaviors
Neuropeptide Y (NPY) is a highly conserved endogenous peptide consisting of 36 amino acids, which is widely distributed in the CNS and PNS of all mammals and acts as a neurohormone and neuromodulator. NPY exerts its biological functions via the interaction with five subtypes of GPCRs
[60][100]. NPY and its receptors are involved in a variety of behaviors, such as food intake, circadian rhythms, chronic pain, the stress response, and anxiety
[61][62][63][64][65][66][101,102,103,104,105,106]. NPY is expressed in many nuclei associated with sleep–wake regulation, such as the amygdala, hypothalamus, hippocampus, periaqueductal gray, LC, and the cerebral cortex
[67][68][69][107,108,109]. Therefore, NPY is thought to affect sleep–wake behaviors
[70][110].
2.6. Substance P Induces Either Sleep or Arousal
Substance P (SP) is a neuropeptide consisting of 11 amino acids
[71][119]. It exerts its functions by binding to neurokinin receptors, particularly neurokinin type 1 receptors (NK-1Rs) and NK-2Rs
[72][120]. NK-1Rs in the CNS are critical for the regulation of affective behaviors, neurochemical responses to stress, and pain transmission
[73][74][121,122]. NK-1Rs are distributed throughout the CNS, including many brain regions that are highly involved in sleep–wake regulation, such as the hypothalamus, brainstem, and cortex
[75][76][123,124]. Therefore, SP affects sleep–wake states.
SP is reported to induce sleep. For example, bilateral microinjection of SP into the VLPO increases NREM sleep in rats
[77][125]. Microinjection of SP into the cerebral cortex enhances the slow-wave activity in mice as well
[78][126]. Furthermore, a previous study has shown that the intracerebroventricular administration of SP conjugated with cholera toxin A subunit can enhance NREM sleep but induce sleep fragmentation
[79][127]. However, on the contrary, the systemic administration of non-nociceptive doses of SP has been reported to increase the duration of wakefulness episodes in mice
[80][128], and to increase the latency of REM sleep and wakefulness in healthy young men
[81][129]. Sergeeva et al. have also found that SP induces arousal by activating histaminergic neurons in the TMN
[82][130].
2.7. Vasoactive Intestinal Peptide Promotes REM Sleep
Vasoactive intestinal peptide (VIP), a peptide consisting of 28 amino acids, is produced in many regions of the human body, including the gut, pancreas, and suprachiasmatic nucleus (SCN)
[83][133]. The SCN is the center of the circadian rhythm, and the circadian rhythm determines sleep–wake states in mammals. Disruption of the circadian rhythm usually causes sleep disorders
[84][134]. VIP, a neurotransmitter expressed by a subset of the SCN neurons, appears to play a critical role in the regulation of the circadian rhythm and sleep–wake behaviors
[85][86][87][135,136,137]. VIPergic neurons project densely throughout the SCN. A previous study revealed that VIP kickout mice showed an 8-h advance of the predicted activity phase with less precision when exposed to constant darkness
[88][138]. This shows that VIP and its receptors are critical for maintaining normal circadian rhythms, which is a significant function of the SCN.
2.8. Other Neuropeptides Associated with Sleep and Wake Regulation
Previous studies have shown that neuropeptides have diverse effects on sleep–wake regulation in rodents and humans.
Here, we summarize the roles of other neuropeptides in sleep–wake regulation. Adrenocorticotropic hormone, cocaine- and amphetamine-regulated transcript, ghrelin, neurotensin, pituitary adenylyl cyclase-activating polypeptide, and somatostatin induce arousal
[89][90][91][92][93][94][146,147,148,149,150,151]. Some of them (neurotensin, pituitary adenylyl cyclase-activating polypeptide, and somatostatin) tend to suppress NREM sleep, but they have a positive impact on REM sleep
[91][92][93][148,149,150]. Other neuropeptides, including brain-derived neurotrophic factor, growth hormone, growth hormone-releasing hormone, interleukin 1 beta, leptin, melanocyte-stimulating hormone, neuropeptide B, opioid peptides, and tumor necrosis factor, can promote NREM sleep
[95][96][97][98][99][100][101][152,153,154,155,156,157,158]. Cholecystokinin has been shown to play a complex role in the regulation of sleep–wake behaviors, because it induces both NREM sleep and wakefulness
[97][154]. Recent studies have also suggested that both cholecystokinin and transforming growth factor alpha can potentially inhibit the transition from sleep to wakefulness
[102][159].