1. Taxonomy of Citrus Canker Bacterium
Citrus canker, also known as Asiatic CC, was initially reported on in the United States of America in the early 1900s following an outbreak in numerous southeastern states
[1][26]. In 1914, Hasse received samples from Florida, Texas, and Mississippi, and was able to isolate the bacterium
[2][27]. After completing characterization and pathogenicity tests, Hasse called the bacterium
Pseudomonas citri [2][27]. Since then, the bacterium has been classified into several genera, including
Bacterium,
Phytomonas, and, finally,
Xanthomonas citri in 1939
[3][4][5][8,28,29].
Xanthomonas genus consists of 27 phytopathogens that cause critical diseases in ornamental plants and other crops
[3][8]. The genus has a broad range of 68 host families, over 240 genera and 140 different pathovars
[6][30]. The genus
Xanthomonas can infect more than 350 species, including 268 dicots and 124 monocots, including grains, fruits, nuts, and plants belonging to
Brassicaceae and
Solanaceae families
[3][8]. The strains of
Xanthomonas citri have been assigned to the A strain within this species to show that they are linked to Asiatic CC
[3][8]. Two more CC-producing
Xanthomonads were discovered in the 1970s and were first classified as group C strains, which induce canker lesions solely in key lime (
Citrus aurantifolia), and group B strains, which have a broader host range
[7][8][31,32]. CC bacterium continued as
X. citri until 1978; in the same year, 1978, Dye placed
X. citri in
X. campestris pv.
citri to uphold
citri at the infra subspecific level
[9][33]. CC bacterium was again reassigned the title of
X. citri by
[10][34], while the B and C strains were placed in
X. campestris pv.
aurantifolii. Reference
[11][35] disagreed with previous
onrese
arch and argued that more work was needed to place CC strains in
X. citri and suggested A, B, and C strains continued as
X. campestris pv.
citri; then
[12][36] performed research using DNA–DNA hybridization (DDH) based on renaturation rates with a diverse array of
Xanthomonas strains, recommending strain A to
X. axonopodis pv.
citri and B and C to
X. axonopodis pv.
aurantifolii, respectively
[3][13][7,8].
FThe research o
rn CC bacterium taxonomy continued and, Ref.
[14][37] again, using the S1 nuclease DNA–DNA hybridization technique, the researchers recommended
X. axonopodis pv.
citri strains in
X. smithii subsp.
smithii and the
X. axonopodis pv.
aurantifolii strains in
X. fuscans subsp.
aurantifolii, although the placement of strains in
X. smithii after due deliberations was later considered illegitimate and was agreed upon by the previous legitimate proposal by
[10][34]. Hence, the
author
s of reference
[15][38] published an erratum “Emended classification of
Xanthomonad pathogens on citrus” in systematic and microbiology and recommended the placement of strains in
X. citri. Finally,
the authors of reference
[16][39] formally validated
X. citri. as
X. citri. subsp.
citri. Reference
[17][40] proposed important modifications to the taxonomy of
Xanthomonads, including within
X. citri, recommending the addition of several pathovars within
X. axonopodis, as well as the placement of members of
X. fuscans, into
X. citri, using a polyphasic approach that included a multilocus sequence analysis (MLSA), a DDH calculation of whole-genome average nucleotide identity values, and phenotypic analyses. As a result, it has been suggested that
X. fuscans subsp.
aurantifolii be transferred to
X. citri as
X. citri pv.
aurantifolii [17][40].
ItThe waauthors submitted
rtheir recommendations for these adjustments to the
International Journal of Systematic and Evolutionary Microbiology, which were accepted; from now on, the prokaryotic names
X. citri subsp.
citri (
XCC) and
X. fuscans subsp.
aurantifolii (
XFA) will be used in nomenclature for bacteria that cause CC
[17][40]. The bacterium was gram-negative, rod-shaped, and polar flagella. In contrast, colonies of bacterium showed yellow colors on petri plates due to the presence of a carotenoid pigment called Xanthomonadin. Because of exopolysaccharide (EPS), it is known as xanthan, showing a glossy appearance, invitro
[17][40]. The classification of bacterium consists of kingdom: Prokaryote, phylum: proteo-bacteria, class: Gamma-proteobacteria, order:
Xanthomonadales, family:
Xanthomonadaceae, genus:
Xanthomonas, specie:
citri, and subsp.: citri
[13][7].
2. Phylogenetically Distinct Groups of CC
X. citri subsp.
citri and
X. citri subsp.
aurantifolii have been further divided into sub-groups based on their significant differences in the host range, which is also a reason for true pathogenic variants
[10][18][34,45]. It was reported that the division of these groups based on citrus host type and symptoms was made on bacterial isolation on various nutrient media
[7][31].
3.1. Asiatic Citrus Canker Strains
The most important and widespread pathovar is the Asiatic-canker (also named cancrosis-A or true-canker),
X. citri subsp.
citri A strain is the most virulent and has a wide host range, including cultivars of citrus
[19][46]. South-West Asian strains
X. citri subsp.
citri A are relatively less widespread
[13][7]. Most recently, in Florida, at one location, a third pathogenic strain was found, which was designated Aw, apparently of Asiatic origin
[20][47].
3.2. South-American Canker Strains
There are two types of South American canker strains which causes the same symptoms on the susceptible host as those produced by
X. citri subsp.
citri A strains, but all strains of South America have narrow host ranges
[20][47].
X. citri subsp.
aurantifolii B strain, also referred to as false canker or cancrosis B, has a more restricted host range and is found to primarily infect lemons and limes
[8][32]. The Bstrain (
XAUB as an acronym, XAC pathotype B;
XAC-B) first appeared in Argentina in 1923 and it eventually extended to nearby Uruguay and Paraguay
[8][9][32,33]. The B strain generally causes severe infections on lemon fruits (
Citrus limon), limes (
C. aurantiifolia), sour oranges (
C. aurantium), but seldom on sweet oranges (
C. sinensis) and pummelo fruit (
C. maxima). Moreover, this strain does not infect grapefruit (
C. paradisi)
[6][8][30,32]. Hence, the Bstrain is not present in nature longer. Mexican lime cancrosis, CBC-C (
XACtype C;
XAC-C) or the Cstrain (
XAUC as an acronym) was discovered in 1963 and present only in São Paulo, Brazil, where it just infects the Mexican lime
[21][48]. The B and C strains are currently classified as
X. axonopodis subsp.
aurantifolii and produce many similar symptoms on the host as produced by the canker A strain
[22][23][49,50]. The strains
XAC,
XAUB, and
XAUC were compared and analyzed phenotypically and phylogenetically; all three strains were shown to have polar flagella with noticeable motility when cultured in semi-solid media
[24][51]. In the presence of maltose and aspartic acid, only XAC can grow and hydrolyze pectate and gel
[25][52]. Polyclonal antisera were prepared against
XAC, but
XAUB and
XAUC showed little or no affinity to antisera. In contrast,
XAC is susceptible to CP1 and CP2 bacteriophages, and
XAUB and
XAUC are not affected by these bacteriophages
[26][53]. It was observed in culture media that
XAUB has fastidious growth;
XAC and
XAUC both grow well in nutrient-agar (NA) and tryptophan–sucrose agar media. Moreover, these three strains show good growth in glutamic-acid rich media. A molecular analysis confirmed that
XAUB and
XAUC are more interlinked with one another than
XAC [27][28][29][54,55,56]. In contrast, data obtained from physiological tests, i.e., phage-typing
[30][57] total protein profiles after SDS-PAGE, DNA–DNA solution hybridizations
[29][31][56,58], plasmid–DNA fingerprints
[32][59], plasmid-based hybridization probes
[32][59], PCR assay
[33][60], DNA fragment sequence of gene
hrp, restriction enzymes to analyze DNA fragments
[34][61] confirm these conclusions. Furthermore, a gene required to cause CBC symptoms is the
pthA gene, which was obtained from the
XAC-A strain
[35][36][62,63]. Total DNA hybridization with a
pthA fragment revealed several profiles among
XAC-A,
XAUB, and
XAUC; no hybridization with strains of
X. axonopodis pv.
citrumelo was observed
[37][10].
Provisionally, two more CC strains were classified, named D and E strains, but now they do not exist or are categorized differently
[38][64]. The D strain, which is also referred to as bacteriosis, induces disease in limes in Colima (Mexico), but its etiology is not confirmed yet
[39][40][65,66]. This disease causes typical leaves and twig lesions, but no symptoms are observed on key Mexican lime fruit
[41][67]. In this area, the suspected citrus pathogen no longer exists. It is now believed that the disease was caused by
Alternaria limicola [42][43][68,69]. The second pathogen was the E strain, previously identified as a citrus canker in a Florida nursery. The disease is ‘called’ a bacterial spot of citrus produced by
X. axonopodis subsp.
citrumelo [10][23][44][34,50,70]. It can be distinguished based on these studies that three groups of
X. axonopodis strains, i.e., Astrain, Bstrain (including C and D strains), cause citrus cankers
[23][50] and, notably, these strains have controversial taxonomy
[45][71]. This interpretation was confirmed when the
Xanthomonas genus was reclassified based on DNA–DNA hybridization and metabolic activity studies
[3][13][37][7,8,10]. Moreover,
Xanthomonas, causing diseases on citrus, was transferred from
X. campestris to
X. axonopodisspecies. Perhaps now, pathotype A, pathotypes B and C, and the CBS strains of
X. axonopodis, are named
X. axonopodis subsp.
citri,
X. axonopodis subsp.
aurantifolii, and
X axonopodis subsp.
citrumelo, respectively, but the subcommittee on the taxonomy of plant pathogenic bacteria did not support this proposal
[3][13][45][7,8,71].
3. Symptomatology
Canker symptoms are observed in all aerial parts of the plant
[46][72]. They are characterized mainly by the formation of erumpent, corky, and raised pustules on the surface of leaves, fruits, and twigs, which serve as sources of bacterial inoculums
[46][72]. Defoliation and fruit drop are also observed as plant responses to the infection
[47][48][73,74]. Notably,
Xanthomonas citri can survive in such plant debris for two months
[49][75]. Severe symptoms are produced on trifoliate oranges, grapefruit, Mexican lime, and some sweet oranges; however, the actual host range depends primarily on a strain of citrus canker
[48][74]. Generally, the susceptibility of young tissues to the citrus canker is more than mature tissues as there is a period of vulnerability in each flush around three times a year
[50][76] (
Figure 12).
Figure 12. (A) Raised, corky, and sunken lesions on the upper side of the leaf. (B) Lesions on the lower side of the leaf. (C) Initial lesions on the lower surface of the leaf. (D) Canker symptoms on the fruit.
3.1. Leaf Lesions
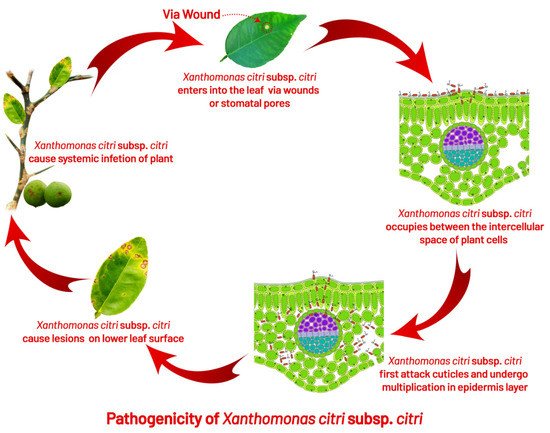
CC bacterium naturally penetrates the host tissues through stomata
[51][77], hydathodes, lenticels, or wounds
[52][78]. Citrus canker disease symptoms first appear as tan, brown, or grey-oily circular lesions, 2 to 10 mm in size, depending on the susceptibility of the host, the number of cycles of the infection, and optimal environmental conditions, i.e., the presence of water film and 20 to 30 °C temperature; canker protrudes from both surfaces of leaf tissue around 4–7 days after inoculation
[52][78]. Symptoms might appear after more than 60 days under optimum conditions
[53][54][79,80]. As the disease advances, host cell expansion (hypertrophy) and cell division (hyperplasia) occur, due to which the lesions become visible from small water-soaked spots and are surrounded by a yellow halo, which turns into slightly raised blister-like lesions and can be viewed with transmitted light
[48][54][74,80]. The hyperplastic mesophyll tissue is an essential diagnostic symptom of the disease characterized by the formation of the canker due to rupturing the epidermis
[52][78] and it releases abundant
X. citri subsp.
citri on the leaves. These lesions are elevated, are ‘corky’ in leaves, stems, and fruits, and then become dark and thick into the distinctive citrus canker under dry conditions
[47][73]. A wound on the leaves or fruits or an injury by the Asiatic citrus leaf miner (
Phyllocnistis citrella) significantly increases symptom severity
[55][56][57][81,82,83].
3.2. Fruit Lesions
Fruits are susceptible for 90–120 days when they grow between 2.0 and 6.0 mm in diameter, depending on citrus species
[51][77]. The lesions in the early stages look similar to large oily glands on the peel and become progressively dark and corky in texture, usually circular, and may occur individually or in groups, leading to premature fruit fall
[58][84].
3.3. Twig Lesions
Twig lesions generally occur when leaves and fruits pass through one or more cycles of infection. Similar symptoms are produced on both twigs and fruits; twig lesions are not surrounded by chlorosis (but fruit lesions do)
[59][24]. Citrus canker is endemic, the inoculum spreads by twig lesions on young shoots, and
X. axonopodis subsp.
citri. survival is prolonged in these areas; lesions with raised corky patches may persist for many years until girdling infections do not kill the twigs
[49][75]. The highest susceptibility of citrus to
X. axonopodis subsp.
citri infection is during the last half of the growth development phase in all of the above ground citrus tissues
[60][85]. Lesion incidence is seasonal, but sometimes severe precipitation and high temperatures coincide with periods of flush growth
[58][84]. As leaves, stems, and fruits are fully grown, they become resistant to infection; once leaves are expanded between 50 and 80%, they become most susceptible
[61][86]. New flushes, tender leaves, and stems are more likely to be vulnerable to citrus cankers than fully grown citrus
[60][85]. When a pathogen severely attacks the host, it leads to defoliation, dieback, early fruit drop, and tree decline; hence, infected fruit is less valuable or unmarketable
[62][63][87,88].
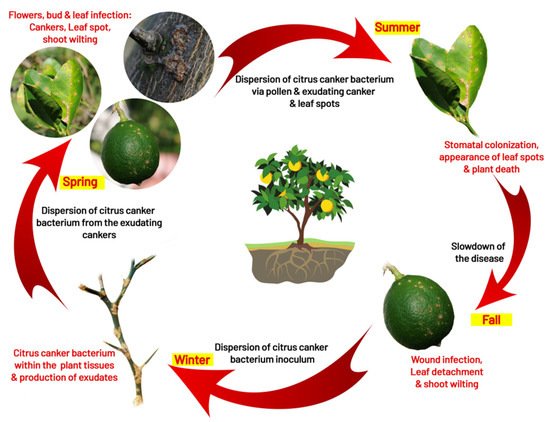
4. Disease Cycle and Epidemiology
4.1. Infection
The bacteria penetrate the host by disrupting the leaf epidermis, inducing cell hyperplasia, and colonizing the apoplast
[64][89]. Under optimum conditions, the pathogen multiplies 3 to 4 log units per lesion; for further disease development, bacterial cells may emerge from stomata openings to provide inoculum within five days
[49][75]. For successful infection and lesion formation, free moisture for 20 min is required for the bacterial cells to ooze out from the lesion. As a result of water congestion, one to two bacterial cells are released from stomatal openings during inoculation
[51][65][77,90]. After the initiation of growth of the host, almost all infections take place on stems and leaves within the first six weeks, while the first 90 days after petals falling is the most crucial time period for fruit infection
[65][90]. Small and unnoticeable pustules are formed due to infections after this time period
[37][10]. It has been reported that fruits are more susceptible to disease than leaves; hence, observations have been made that lesions of different sizes can be found on the rind of the same fruit during the infection of the bacterium
[66][91] (
Figure 3).
Figure 23.
Dispersion of citrus canker bacterium in orchards.
4.2. Survival
The main inoculum sources are branches, leaves, and twigs infected with cankers
[58][84]. The disease is primarily carried in cankers on twigs and branches from one season to another, serving as a primary inoculum
[37][10]. In leaf and fruit lesions, the bacteria remain alive until they fall; because the affected leaves fall off early, they may not act as the primary inoculum source
[49][75]. Still, it was reported that, in infected leaves, the bacterium survived up to six months
[67][92]. Reference
[68][93] found that the bacterium survived for over 6months in the infected leaves, for 52 days in sterilized soil, and only 9 days in unsterilized soils, respectively. It was also observed that the organism could survive for 11–12 days under desiccation at 30 °C
[68][93]. On citrus hosts, the bacterium survives epiphytically with a lower population without developing the symptoms, combined with non-citrus weeds, grass host, and soil
[69][70][71][94,95,96]. However, in the absence of plant tissue or debris, the saprophytic existence of soil pathogens has not been observed
[68][70][93,95]. The survival capability of the pathogen in subtropical soil is very limited
[72][97], and bacterial inoculum dies within 24 to 72 h on different inert surfaces, such as cloths, metals, plastics, and processed wood in both sunlight and shade
[66][91]. Due to antagonisms and competition with saprophytic microorganisms, the bacterial population decreases to an undetectable level 1–2 months after leaves or fruits fall to the ground
[66][91]. In Japan and Brazil, it has been reported that
X.
axonopodis subsp.
citri may survive on non-host plant material and in the root zone of certain weeds under eradicated diseased trees for a few weeks
[73][98] (
Figure 34).
Figure 34.
How canker bacterium initiates local infection into leaves, twigs, and fruits.
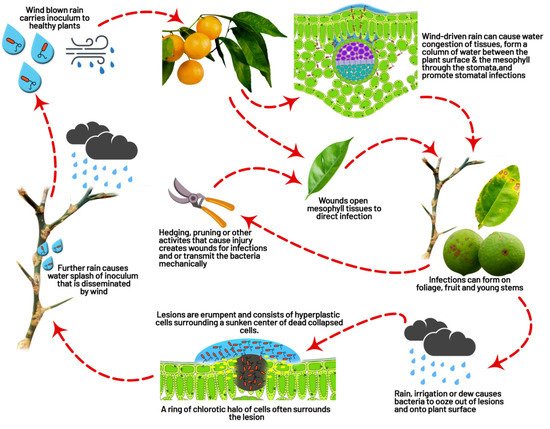
4.3. Dispersal
Under natural conditions, rainfall splashes and rainfall associated with wind are reasons for the short-distance spread of disease. Still, dissemination to long-distances between geographical regions is mainly expected to occur through infected plant material
[59][24]. During severe storms, such as tornadoes, disease spread takes up to 10 to 15 km
[65][90]. Dispersal of diseases have been further explored with model-based data on wind direction and threshold parameters, which showed wind speed eight m/s and rainfall of nearly 0.32 cm/h helped the insects, such as
P. citrella, and blowing sand penetrates the bacteria through stomatal pores or injuries caused by thorns
[74][99]. The major reason for the dispersal of bacteria to average distance is the wind-driven rain. In Argentina, the wind-blown rain dispersed the bacteria from infected trees up to a distance of 32 m
[75][100]. A drop of rainwater may contain up to 105 to 108 cfu/mL of bacteria
[54][75][80,100]. Under globalization, the transmission of bacteria from one region to another through frequent communication and transportation increases the risk of infecting citrus farming in free areas of disease
[76][101] (
Figure 45).
Figure 45.
Cellular interaction of
Xanthomonas citri
subsp.
citri
with the host and how it expresses symptoms on host plant parts.
4.4. Role of Insect (Leaf Miner Interaction)
The leaf miner plays an important role in the spread of citrus canker, but it has not yet been reported as a disease vector
[55][81]. In earlier 1994, the distribution of citrus leaf miners was restricted to southeast and southwest Asia, while it spread after the mid-1990s to most of the world’s major citrus areas
[55][81]. It was first reported in Florida and Brazil in 1993 and 1996
[77][102]. The feeding activities of the citrus leaf miner provide bacterial infections to the host in three ways: (1) wind-blown rain disseminates the bacteria, contacts the surface of the leaf; the leaf miner tears the cuticle and opens the mesophyll of the leaf, providing direct bacterial infection; (2) the leaf miner injuries are cured more slowly than mechanical injuries, which allow for longer exposure to bacterial infections; (3) leaf miner larvae may become contaminated with bacteria and carry it to the feeding galleries, where feeding activities lead to an increase in mesophyll cells infection
[74][78][99,103]. Trees with leaf miner injuries remain susceptible for 7–14 days, compared with only 24 h for wind, thorns, or pruning injuries
[79][104]. The prevalence of citrus canker increases in Brazil and Florida due to leaf miner injuries
[48][65][79][74,90,104]. Still, it is believed that the host can tolerate some loss of leaf area without yield being affected because of leaf miner damage (up to 10%)
[80][105]; there
iare reports
that loss of 16–23% of leaf areas can lead to significant yield loss
[81][106].