Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Camila Xu and Version 1 by Gaosi Xu.
由严重急性呼吸系统综合征冠状病毒2(SARS-CThe coroV-2)引起的2019年冠状病毒病(navirus disease 2019 (COVID-19)大流行对全球经济和健康造成了异常的挫折。疫苗接种是显著减少) pandemic, caused by the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), has caused an exceptional setback to the global economy and health. Vaccination is one of the most effective interventions to markedly decrease severe illness and death from COVID-19重症和死亡的最有效干预措施之一。.
- acute kidney disease
- acute kidney injury
- COVID-19
1. Introduction
With the ongoing coronavirus disease 2019 (COVID-19) pandemic and the emergence of new variants of severe acute respiratory syndrome coronavirus type 2 (SARS-CoV-2), the rapid development of effective and safe preventive vaccines is urgently required to control disease outbreaks [1,2][1][2]. Over the past 2 years, hundreds of COVID-19 vaccine candidates have been developed, tested, and finally rolled out, including protein-based vaccines (Novavax), inactivated vaccines (Sinovac Life Science), viral vector vaccines (Janssen, Oxford-AstraZeneca), and mRNA vaccines (Pfizer/BioNtech, Moderna, CureVac) (Figure 1) [2,3][2][3]. Among them, mRNA-based drugs are new but not unknown [4]. mRNA vaccines deliver transgenic mRNA through lipid nanoparticles, which act as carriers. Once injected, the mRNA is translated into the target protein in vivo, resulting in a strong immune response, and a 2-dose regimen confers 95% protection against COVID-19 [5]. To date, large phase III and IV trials have found these vaccines to have a good safety profile, with few serious reactions [3,6,7,8,9][3][6][7][8][9]. Common short-term adverse events include local injection site reactions, fever, fatigue, generalized pain, and headache [6,10][6][10].
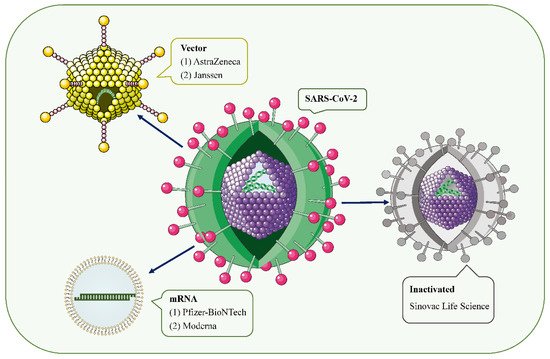
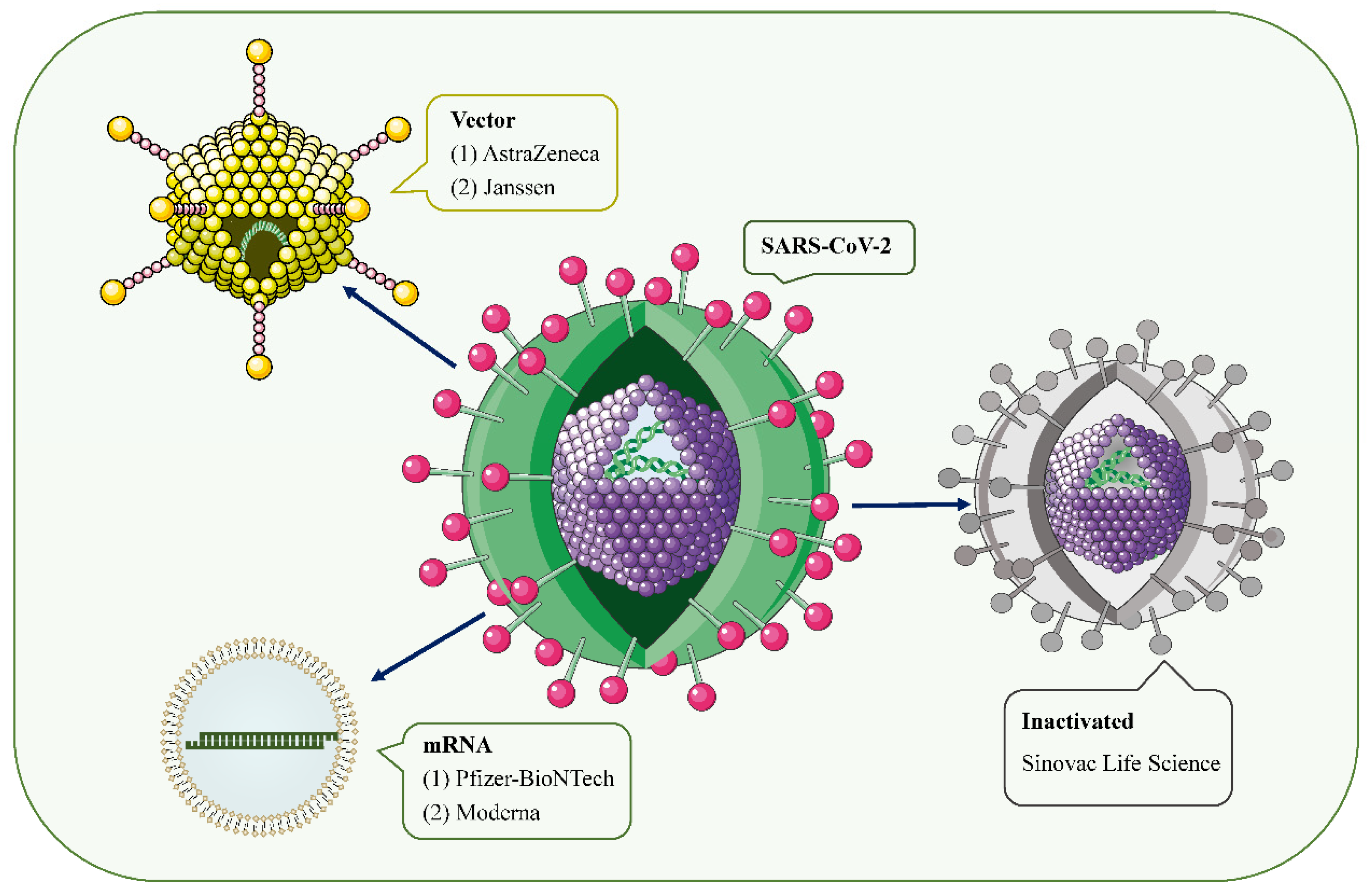 Figure 1. SARS-CoV-2 and the main types of vaccines that may trigger AKD. SARS-CoV-2 is a positive-sense single-stranded RNA virus with a lipid bilayer consisting of the spike S protein and membrane and envelope proteins. mRNA vaccines deliver transgenic mRNA through lipid nanoparticles as carriers. Viral vector vaccines utilize adenovirus and integrate genetic material from SARS-CoV-2 into its genome. Inactivated vaccines involve SARS-CoV-2 that has been killed by physical or chemical means.
Figure 1. SARS-CoV-2 and the main types of vaccines that may trigger AKD. SARS-CoV-2 is a positive-sense single-stranded RNA virus with a lipid bilayer consisting of the spike S protein and membrane and envelope proteins. mRNA vaccines deliver transgenic mRNA through lipid nanoparticles as carriers. Viral vector vaccines utilize adenovirus and integrate genetic material from SARS-CoV-2 into its genome. Inactivated vaccines involve SARS-CoV-2 that has been killed by physical or chemical means.However, since mass vaccination, there have been a few case reports of acute kidney injury (AKI), acute kidney disease (AKD), proteinuria, edema, gross hematuria, and other renal side effects requiring hospitalization after COVID-19 vaccinations [11]. Serum creatinine (Scr) levels and proteinuria recovered within 3 months of treatment in most patients. The vast majority of cases occurred after mRNA vaccine and adenoviral vector injection, and a few cases of glomerulonephritis associated with inactivated virus vaccines have also been reported.
2. Inducing AKD through COVID-19 Vaccine: Hypotheses
2.1. Podocyte Damage
The temporal association between intramuscular vaccination and the development of MCD speculates that a cell-mediated immune response may be a trigger for podocyte injury [20,53][12][13]. All 12 patients with MCD reported in the literature were over 60 years of age, developed AKD within 2 weeks of vaccination, and steroids appeared to be effective in achieving rapid remission (Table 1). Typically, following vaccination, the vaccine’s antigens are taken up by dendritic cells and then presented to T cell receptors on naive T cells [54][14]. This leads to the activation of antigen-specific effector T cells, peaking 7 to 14 days after vaccination [55][15]. Studies have also confirmed that during viral infection, cellular immune responses can be observed within about 1 week after infection, but T cell activation can occur 2–3 days earlier [56,57][16][17]. This answers the question of whether it is reasonable for a COVID-19 vaccine to elicit a cell-mediated response 3–4 days after administration. Although the exact pathogenesis of MCD remains unclear, podocyte damage caused by circulating factors released by activated T lymphocytes appears to be decisive (Figure 2) [58,59][18][19]. During active stages of MCD, T cell subsets are imbalanced, and circulating CD8+ suppresses the prevalence of T cells, which is exacerbated by cytokine-induced damage [60][20]. Compared with conventional vaccines, mRNA vaccines are expected to provoke higher antibody responses and stronger CD8+ T and CD4+ T cell reactions, including higher chemokine and cytokine production [61,62][21][22]. The resulting irregular permeability factors can alter glomerular permeability and lead to marked proteinuria and kidney injury [53][13].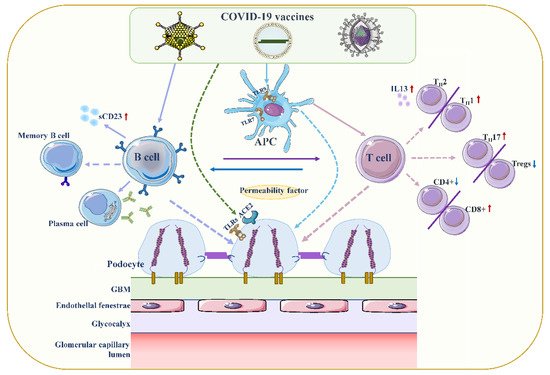
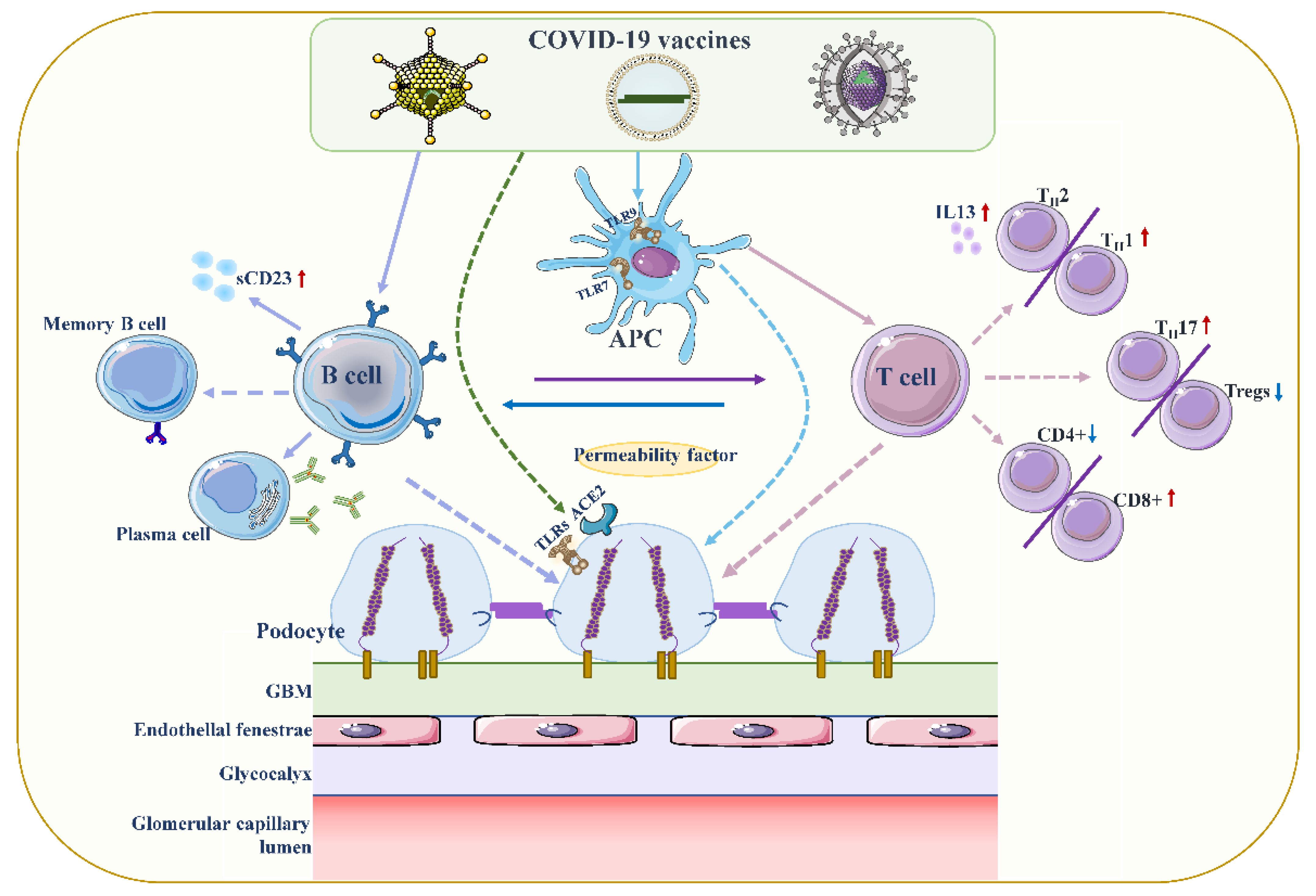 Figure 2. Proposed mechanisms of podocyte injury caused by COVID19 vaccination. Vaccination stimulates antigen-presenting cells (APCs) and B cells, which in turn activate T cells through antigen presentation and cytokine production. A decrease in CD4+ T helper (Th) cells is associated with the prevalence of CD8+ cytotoxic T cells, and an imbalance between Th2 and Th1 cells is associated with an increase in Th2-specific interleukin-13 (IL-13) production, and Th17. In contrast to increased cellular activity, the frequency and function of regulatory T cells (Tregs) decreased. Permeability proteins, such as cytokines and autoantibodies, can directly affect podocytes, leading to loss of foot processes and disruption of the glomerular permeability barrier. In addition, the vaccine can also affect podocytes through specific toll-like receptors (TLRs), and angiotensin conversion enzyme 2 (ACE2). The figure refers to the pathogenesis of minimal change disease by Vivarelli et al [53][13].
Figure 2. Proposed mechanisms of podocyte injury caused by COVID19 vaccination. Vaccination stimulates antigen-presenting cells (APCs) and B cells, which in turn activate T cells through antigen presentation and cytokine production. A decrease in CD4+ T helper (Th) cells is associated with the prevalence of CD8+ cytotoxic T cells, and an imbalance between Th2 and Th1 cells is associated with an increase in Th2-specific interleukin-13 (IL-13) production, and Th17. In contrast to increased cellular activity, the frequency and function of regulatory T cells (Tregs) decreased. Permeability proteins, such as cytokines and autoantibodies, can directly affect podocytes, leading to loss of foot processes and disruption of the glomerular permeability barrier. In addition, the vaccine can also affect podocytes through specific toll-like receptors (TLRs), and angiotensin conversion enzyme 2 (ACE2). The figure refers to the pathogenesis of minimal change disease by Vivarelli et al [53][13].2.2. Increased Production of Anti-Neutrophil Cytoplasmic Autoantibodies (ANCAs)
基于病毒性Influenza and rabies vaccines based on viral mRNA的流感和狂犬病疫苗已被描述可能导致ANCA升高,从而导致ANCA相关血管炎的发展[67]。此外,证实在用核糖核酸酶治疗疫苗后,s have been described to possibly lead to an increase in ANCA, contributing to the development of ANCA-associated vasculitis [28]. Moreover, it was confirmed that the ANCA反应显着降低。科学家发现,在 response was significantly reduced after the treatment of vaccinees with ribonuclease. Scientists have found that in the context of COVID-19的背景下,宿主对病毒RNA的反应可直接引起ANCA相关血管炎(AAV)和自身免疫反应[68,69,70]。与初次免疫相比,, a host response to viral RNA can directly cause ANCA-associated vasculitis (AAV) and an autoimmune response [29][30][31]. COVID-19 mRNA疫苗接种在第二次加强免疫后诱导先天免疫系统的反应更强[71]。第二次接种 vaccination induced a stronger response of the innate immune system after the second booster compared with primary immunization [32]. The heightened innate immune response observed after the second vaccination with BNT162b2 mRNA疫苗后观察到的先天免疫应答增强可能是 vaccine may be an inducer of MPO-ANCA和PR3自身抗体的诱导剂[34]。 and PR3 autoantibodies [33]. Tolll样受体(TLR)可以在白细胞膜上表达,并在炎症反应,识别病毒抗原和促进免疫系统激活中起重要作用。在AAV中,主要收费样受体2(TLR2)和收费样受体9(TLR9)激活可引起自身免疫[72]。有趣的是,-like receptors (TLRs) can be expressed on leukocyte membranes and play an important role in inflammatory responses, recognizing viral antigens and promoting immune system activation. In AAV, major toll-like receptor 2 (TLR2) and toll-like receptor 9 (TLR9) activation can provoke autoimmunity [34]. Interestingly, Kumar等人提出, et al. suggested that TLR2是由免疫显性细胞毒性T淋巴细胞(CTL)对 was activated by a robust and specific immune response of immunodominant cytotoxic T-lymphocyte (CTL) to the spike glycoprotein of SARS-CoV2(也由 (also produced by the COVID-19疫苗产生的)的刺突糖蛋白的强健和特异性免疫反应激活的[73]。信使 vaccine) [35]. Messenger RNA疫苗由于其 vaccines could act as both antigen and adjuvant due to their intrinsic immunostimulatory properties of RNA的内在免疫刺激特性,可以同时作为抗原和佐剂;因此,它们可以通过内体TLR和胞质炎症小体成分来识别[64]。因此,与非; thus, they can be recognized by endosomal TLRs and cytosolic inflammasome comRNponents [25]. Therefore, the occurrence of AA疫苗接种相比,在V in the context of COVID-19 mRNA背景下AAV的发生具有高度相关性,但需要进一步的实验来验证自身免疫与 is highly relevant compared with non-mRNA vaccinations, but further experiments are required to verify the mechanism of the link between autoimmunity and a COVID-19疫苗之间联系的机制。 vaccine.疫苗诱导的血栓性血小板减少症(VITT)
2.3. Vaccine-Induced Thrombotic Thrombocytopenia (VITT)
一些学者推测,抗磷脂抗体(Some scholars have speculated that antiphospholipid antibodies (APL)可能是s) may be part of the cause of thrombosis after COVID-19疫苗接种后血栓形成的部分原因,因为它会触发与APLs产生相关的 vaccination, by triggering the type I interferon response associated with APLs’ production [36][37]. I型干扰素反应[74,75]。它通过抑制蛋白t binds directly to platelets by inhibiting the anticoagulant pathway of protein C的抗凝血途径直接与血小板结合,触发凝血级联反应,并且似乎与涉及补体级联反应的免疫应答的异常激活有关[74]。据报道,在给予腺病毒基因转移载体后,血小板减少和血小板活化[76]。血小板减少症也发生在一些反义寡核苷酸治疗后[77]。基于上述背景,另一种假设推测,由血管性血友病因子(, triggers the coagulation cascade, and appears to be associated with abnormal activation of immune responses involving the complement cascade [36]. Thrombocytopenia and platelet activation have been reported following the administration of adenoviral gene transfer vectors [38]. Thrombocytopenia also occurred after treatment with some anti-sense oligonucleotides [39]. Based on the above background, another hypothesis speculates that the activation of platelets by adenovirus-platelet-leukocyte complexes, mediated by von Willebrand factor (VWF)和) and P-选择素介导的腺病毒selectin, may lead to accelerated clearance of platelets in the liver [37][40]. However, the virus in viral vector vaccines is replication-血小板-白细胞复合物活化血小板可能导致肝脏血小板清除加速[75,78]。 然而,病毒载体疫苗中的病毒是不可复制的,并且循环病毒在接种疫苗后incomparable and the circulating virus disappears 7-–14日消失,因此病毒定位到中枢神经系统和消化系统以引起血栓形成的可能性不大[79]。此外, days after vaccination, so the viral localization to the central nervous system and digestive system causing thrombosis is unlikely [41]. In addition, Greinacher等人提出, et al. suggested that the rare occurrence of VITT的罕见发生是由血小板因子4(PF4)依赖性血小板活化抗体介导的,而血小板激活抗体又通过其Fcγ受体刺激血小板[80,81]。含有 was mediated by platelet factor 4 (PF4)-dependent platelet-activating antibodies, which in turn stimulate platelets via their Fcγ receptors [42][43]. Immune complexes containing PF4的免疫复合物可以被 can be recognized by C1q识别,C1q与IgG分子的Fc部分结合。这导致C3活化,补体反应的扩大以及下游促炎介质和效应子的产生,最终导致血栓炎症增强。, which binds to the Fc portion of IgG molecules. This results in C3 activation, expansion of the complement response, and production of downstream proinflammatory mediators and effectors, ultimately leading to enhanced thrombus inflammation.2.4. 直接诱导肌炎
2.4. Direct Induction of Myositis
一例既往病例报告称,一例患者在接种A previous case reported that a patient who presented with profound left upper arm pain after COVID-19 mRNA疫苗后出现严重左上臂疼痛,血清肌酸激酶浓度升高,提示骨骼肌损伤和炎症(肌炎)[82]。还有证据表明,疫苗接种后患者的肾活检显示大量横纹肌溶解诱导的肌红蛋白投射,这可能导致肾功能恶化[34]。 vaccination had an increased serum creatine kinase concentration, indicating skeletal muscle damage and inflammation (myositis) [44]. There is also evidence of renal biopsies from post-vaccination patients showing massive rhabdomyolysis-induced myoglobin casting, which may contribute to worsening renal function [33].References
- Sahin, U.; Muik, A.; Vogler, I.; Derhovanessian, E.; Kranz, L.M.; Vormehr, M.; Quandt, J.; Bidmon, N.; Ulges, A.; Baum, A.; et al. BNT162b2 vaccine induces neutralizing antibodies and poly-specific T cells in humans. Nature 2021, 595, 572–577.
- Sharma, O.; Sultan, A.A.; Ding, H.; Triggle, C.R. A Review of the Progress and Challenges of Developing a Vaccine for COVID-19. Front. Immunol. 2020, 11, 585354.
- Fiolet, T.; Kherabi, Y.; MacDonald, C.J.; Ghosn, J.; Peiffer-Smadja, N. Comparing COVID-19 vaccines for their characteristics, efficacy and effectiveness against SARS-CoV-2 and variants of concern: A narrative review. Clin. Microbiol. Infect. 2021, 28, 202–221.
- Wolff, J.A.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L. Direct gene transfer into mouse muscle in vivo. Science 1990, 247 Pt 1, 1465–1468.
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615.
- Thomas, S.J.; Moreira, E.D., Jr.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Polack, F.P.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine through 6 Months. N. Engl. J. Med. 2021, 385, 1761–1773.
- Emary, K.R.W.; Golubchik, T.; Aley, P.K.; Ariani, C.V.; Angus, B.; Bibi, S.; Blane, B.; Bonsall, D.; Cicconi, P.; Charlton, S.; et al. Efficacy of ChAdOx1 nCoV-19 (AZD1222) vaccine against SARS-CoV-2 variant of concern 202012/01 (B.1.1.7). an exploratory analysis of a randomised controlled trial. Lancet 2021, 397, 1351–1362.
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2. an interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111.
- Costa Clemens, S.A.; Weckx, L.; Clemens, R.; Almeida Mendes, A.V.; Ramos Souza, A.; Silveira, M.B.V.; da Guarda, S.N.F.; de Nobrega, M.M.; de Moraes Pinto, M.I.; Gonzalez, I.G.S.; et al. Heterologous versus homologous COVID-19 booster vaccination in previous recipients of two doses of CoronaVac COVID-19 vaccine in Brazil (RHH-001). a phase 4, non-inferiority, single blind, randomised study. Lancet 2022, 399, 521–529.
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416.
- Klomjit, N.; Alexander, M.P.; Fervenza, F.C.; Zoghby, Z.; Garg, A.; Hogan, M.C.; Nasr, S.H.; Minshar, M.A.; Zand, L. COVID-19 Vaccination and Glomerulonephritis. Kidney Int. Rep. 2021, 6, 2969–2978.
- D’Agati, V.D.; Kudose, S.; Bomback, A.S.; Adamidis, A.; Tartini, A. Minimal change disease and acute kidney injury following the Pfizer-BioNTech COVID-19 vaccine. Kidney Int. 2021, 100, 461–463.
- Vivarelli, M.; Massella, L.; Ruggiero, B.; Emma, F. Minimal Change Disease. Clin. J. Am. Soc. Nephrol. 2017, 12, 332–345.
- Williams, M.A.; Bevan, M.J. Effector and memory CTL differentiation. Annu. Rev. Immunol. 2007, 25, 171–192.
- Mudd, P.A.; Minervina, A.A.; Pogorelyy, M.V.; Turner, J.S.; Kim, W.; Kalaidina, E.; Petersen, J.; Schmitz, A.J.; Lei, T.; Haile, A.; et al. SARS-CoV-2 mRNA vaccination elicits a robust and persistent T follicular helper cell response in humans. Cell 2021, 185, 603–613.e15.
- Miao, H.; Hollenbaugh, J.A.; Zand, M.S.; Holden-Wiltse, J.; Mosmann, T.R.; Perelson, A.S.; Wu, H.; Topham, D.J. Quantifying the early immune response and adaptive immune response kinetics in mice infected with influenza A virus. J. Virol. 2010, 84, 6687–6698.
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880.
- Colucci, M.; Corpetti, G.; Emma, F.; Vivarelli, M. Immunology of idiopathic nephrotic syndrome. Pediatr. Nephrol. 2018, 33, 573–584.
- Mathieson, P.W. Immune dysregulation in minimal change nephropathy. Nephrol. Dial. Transplant. 2003, 18 (Suppl. S6), vi26–vi29.
- Le Berre, L.; Herve, C.; Buzelin, F.; Usal, C.; Soulillou, J.P.; Dantal, J. Renal macrophage activation and Th2 polarization precedes the development of nephrotic syndrome in Buffalo/Mna rats. Kidney Int. 2005, 68, 2079–2090.
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279.
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses. Nature 2020, 586, 594–599.
- Imanishi, T.; Ishihara, C.; Badr Mel, S.; Hashimoto-Tane, A.; Kimura, Y.; Kawai, T.; Takeuchi, O.; Ishii, K.J.; Taniguchi, S.; Noda, T.; et al. Nucleic acid sensing by T cells initiates Th2 cell differentiation. Nat. Commun. 2014, 5, 3566.
- Kobayashi, S.; Fugo, K.; Yamazaki, K.; Terawaki, H. Minimal change disease soon after Pfizer-BioNTech COVID-19 vaccination. Clin. Kidney J. 2021, 14, 2606–2607.
- Teijaro, J.R.; Farber, D.L. COVID-19 vaccines: Modes of immune activation and future challenges. Nat. Rev. Immunol. 2021, 21, 195–197.
- Gupta, R.K.; Bhargava, R.; Shaukat, A.A.; Albert, E.; Leggat, J. Spectrum of podocytopathies in new-onset nephrotic syndrome following COVID-19 disease. a report of 2 cases. BMC Nephrol. 2020, 21, 326.
- Gabarre, P.; Dumas, G.; Dupont, T.; Darmon, M.; Azoulay, E.; Zafrani, L. Acute kidney injury in critically ill patients with COVID-19. Intens. Care Med. 2020, 46, 1339–1348.
- Jeffs, L.S.; Nitschke, J.; Tervaert, J.W.; Peh, C.A.; Hurtado, P.R. Viral RNA in the influenza vaccine may have contributed to the development of ANCA-associated vasculitis in a patient following immunisation. Clin. Rheumatol. 2016, 35, 943–951.
- Uppal, N.N.; Kello, N.; Shah, H.H.; Khanin, Y.; De Oleo, I.R.; Epstein, E.; Sharma, P.; Larsen, C.P.; Bijol, V.; Jhaveri, K.D. De Novo ANCA-Associated Vasculitis With Glomerulonephritis in COVID-19. Kidney Int. Rep. 2020, 5, 2079–2083.
- Vlachoyiannopoulos, P.G.; Magira, E.; Alexopoulos, H.; Jahaj, E.; Theophilopoulou, K.; Kotanidou, A.; Tzioufas, A.G. Autoantibodies related to systemic autoimmune rheumatic diseases in severely ill patients with COVID-19. Ann. Rheum Dis. 2020, 79, 1661–1663.
- Izci Duran, T.; Turkmen, E.; Dilek, M.; Sayarlioglu, H.; Arik, N. ANCA-associated vasculitis after COVID-19. Rheumatol. Int. 2021, 41, 1523–1529.
- Arunachalam, P.S.; Scott, M.K.D.; Hagan, T.; Li, C.; Feng, Y.; Wimmers, F.; Grigoryan, L.; Trisal, M.; Edara, V.V.; Lai, L.; et al. Systems vaccinology of the BNT162b2 mRNA vaccine in humans. Nature 2021, 596, 410–416.
- Hakroush, S.; Tampe, B. Case Report. ANCA-Associated Vasculitis Presenting With Rhabdomyolysis and Pauci-Immune Crescentic Glomerulonephritis After Pfizer-BioNTech COVID-19 mRNA Vaccination. Front. Immunol. 2021, 12, 762006.
- Summers, S.A.; Steinmetz, O.M.; Gan, P.Y.; Ooi, J.D.; Odobasic, D.; Kitching, A.R.; Holdsworth, S.R. Toll-like receptor 2 induces Th17 myeloperoxidase autoimmunity while Toll-like receptor 9 drives Th1 autoimmunity in murine vasculitis. Arthritis Rheum 2011, 63, 1124–1135.
- Kumar, N.; Admane, N.; Kumari, A.; Sood, D.; Grover, S.; Prajapati, V.K.; Chandra, R.; Grover, A. Cytotoxic T-lymphocyte elicited vaccine against SARS-CoV-2 employing immunoinformatics framework. Sci. Rep. 2021, 11, 7653.
- Talotta, R.; Robertson, E.S. Antiphospholipid antibodies and risk of post-COVID-19 vaccination thrombophilia. The straw that breaks the camel’s back? Cytokine Growth Factor. Rev. 2021, 60, 52–60.
- Chen, Y.; Xu, Z.; Wang, P.; Li, X.M.; Shuai, Z.W.; Ye, D.Q.; Pan, H.F. New-onset autoimmune phenomena post-COVID-19 vaccination. Immunology 2021, 165, 386–401.
- Lai, K.Y.; Au, S.Y.; Fong, K.M. Thrombotic Thrombocytopenia after ChAdOx1 nCoV-19 Vaccination. N. Engl. J. Med. 2021, 385, e11.
- Pasin, F.; Calabrese, A.; Pelagatti, L. Immune thrombocytopenia following COVID-19 mRNA vaccine. casuality or causality? Intern. Emerg. Med. 2022, 17, 295–297.
- Othman, M.; Labelle, A.; Mazzetti, I.; Elbatarny, H.S.; Lillicrap, D. Adenovirus-induced thrombocytopenia. the role of von Willebrand factor and P-selectin in mediating accelerated platelet clearance. Blood 2007, 109, 2832–2839.
- Eichinger, S.; Warkentin, T.E.; Greinacher, A. Thrombotic Thrombocytopenia after ChAdOx1 nCoV-19 Vaccination. Reply. N. Engl. J. Med. 2021, 385, e11.
- Greinacher, A.; Thiele, T.; Warkentin, T.E.; Weisser, K.; Kyrle, P.A.; Eichinger, S. Thrombotic Thrombocytopenia after ChAdOx1 nCov-19 Vaccination. N. Engl. J. Med. 2021, 384, 2092–2101.
- Greinacher, A.; Selleng, K.; Mayerle, J.; Palankar, R.; Wesche, J.; Reiche, S.; Aebischer, A.; Warkentin, T.E.; Muenchhoff, M.; Hellmuth, J.C.; et al. Anti-platelet factor 4 antibodies causing VITT do not cross-react with SARS-CoV-2 spike protein. Blood 2021, 138, 1269–1277.
- Theodorou, D.J.; Theodorou, S.J.; Axiotis, A.; Gianniki, M.; Tsifetaki, N. COVID-19 vaccine-related myositis. QJM 2021, 114, 424–425.
More