Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by John Edward Bradshaw and Version 2 by Rita Xu.
Diploid potatoes can be converted from an outbreeding species, in which self-pollination is prevented by a gametophytic self-incompatibility system, into one where self-pollination is possible, either through a dominant self-incompatibility inhibitor gene (
Sli
) or knockout mutations in the incompatibility locus.
- inbreeding depression
- hybrid vigor
- self-pollination
- self-compatibility
- true potato seed
1. Introduction
The potato (Solanum tuberosum) is the world’s fourth most important food crop after maize, rice, and wheat, with 359 million tonnes fresh weight (FW) of tubers produced in 2020 from 16.5 million hectares of land, in 163 countries, resulting in a global average yield of 21.8 t ha−1 (http://faostat.fao.org, accessed on 4 January 2022). Most cultivated potatoes are tetraploids, and their ancestry can be traced back to autotetraploids (2n = 4x = 48) of the diploid cultigen S. tuberosum Stenotomum Group (2n = 2x = 24), which was domesticated from diploid wild species in the highlands of Southern Peru [1], from at least 7000 years before present [2]. Furthermore, most cultivated potatoes are vegetatively propagated and grown from tubers. However, when the International Potato Centre (CIP) was founded in Lima, Peru, in 1971, propagation by true potato seed (TPS) appeared an attractive proposition for the torrid zones of the lowland tropics and subtropics. By 1994, Golmirzaie et al. [3] thought that TPS technology could be the basis of a new green revolution, designed specifically to improve the production and consumption of potatoes in the developing countries of the torrid zone. Later crop maturation and less genetic uniformity could be outweighed by the reduced costs of planting material (TPS), flexibility of planting time, and freedom from tuber-borne diseases [3]. In contrast, when Chilver et al. [4] reviewed on-farm profitability and prospects for TPS, they concluded that widespread geographic adoption was unlikely in the immediate future, but that investment in a small but sustained TPS breeding effort could be justified in both China and India. However, there has been renewed enthusiasm for TPS technology since the advent of diploid F1 hybrid breeding in 2008 [5], as it made possible the production of genetically uniform cultivars. More generally, Jansky et al. [6] and Jansky and Spooner [7] have argued that the next step in the development of potato breeding may be a return to the diploid level to implement an inbred-line-based strategy leading to F1 hybrid cultivars. This would allow potato breeders to reduce the genetic load of undesirable alleles through inbreeding and then combine desirable traits through hybridization, as also pointed out by Bachem et al. [8].
2. Mendel, Darwin, and Fisher
Darwin studied the effects of cross- and self-fertilization in 57 species of flowering plants from 52 genera and 30 families [9][10]. He conducted experiments over 11 years, involving up to 10 successive generations of self-pollination and the growing of offspring in pots in his greenhouse or in rows in his garden. His most important conclusion was that cross-fertilization is generally beneficial and self-fertilization (inbreeding) injurious, and hence the reason for the existence of mechanisms to promote cross-pollination. He did, however, find that some species were tolerant to inbreeding, although crosses between cultivars displayed hybrid vigor. One such species was the garden pea (Pisum sativum), chosen by Mendel for his experiments in plant hybridization that led to his laws of inheritance [10][11]. Mendel’s work was unknown to Darwin, and remained unknown to the world until rediscovered in 1900, translated into English in 1901, and promoted by Bateson [11][12]. In retrospect, the work of Mendel on the mechanism of inheritance, and of Darwin on the mating system, can be viewed as the foundations of scientific plant breeding from the beginning of the 20th century. The world’s four most important food crops have three different mating systems. Wheat (Triticum aestivum) and rice (Oryza sativa) are inbreeding, seed-propagated crops; maize (Zea mays) is an outbreeding, seed-propagated crop; and the potato (S. tuberosum) is an outbreeding, but primarily clonally propagated, tuber crop. It can be argued that, as a consequence, maize breeding has benefited the most, and potato breeding the least, from underpinning genetics research [12][13]. Darwin was particularly interested in the evolutionary advantages and disadvantages of the variation found in breeding and mating systems, and how one system could evolve into another, such as the shift from crossbreeding to inbreeding [9][13][10,14]. Fisher combined the work of Darwin and Mendel into a theory of inbreeding, with the important addition of an explanation for disadvantageous genes (alleles) tending to be recessive [14][15]. Furthermore, he advocated greater use of inbreeding for the practical improvement of domestic plants and animals, given the resounding success of the methods adopted for maize improvement based on inbreeding. Indeed, one of the major achievements in 20th century plant breeding was the production of high yielding, genetically uniform F1 hybrid cultivars of outbreeding crops from genetically variable, open-pollinated ones. Thus, let reusearchers return to the breeding of diploid potatoes, starting with diploid potato germplasm.3. Diploid Germplasm
Andean farmers still cultivate diploid landraces and named cultivars of the Stenotomum Group, the Goniocalyx Subgroup, and the more widely grown Phureja Group, of S. tuberosum [15][16]. Such germplasm is currently proving valuable for the genetic biofortification of potatoes with iron and zinc [16][17][18][17,18,19]. Furthermore, from the 1960s, potato breeders in North America and Europe successfully widened the genetic base of their breeding programs through the selection of long-day-adapted diploid populations of Phureja/Stenotomum Groups from landraces held in genebanks [19][20][21][20,21,22]. Diploid cultivars have been produced from this germplasm such as Mayan Gold, Inca Sun, Inca Dawn, Mayan Queen, Mayan Star, and Mayan Twilight, as part of a program in Scotland [22][23]. Since the 1960s, it has also been possible to reduce tetraploid S. tuberosum to the diploid level through crosses with 2x Phureja Group ‘pollinators’ that induce dihaploid production by a form of parthenogenesis [23][24][24,25]. Subsequently the frequency of dihaploids (also referred to as haploids) has been increased by the use of elite pollinators, such as IVP35, IVP48, IVP101, and PL-4 [25][26][26,27], although the frequency is still low, with reported values of 4.6 to 25.6 dihaploids per 100 berries [27][28]. Busse et al. [27][28] have described a high throughput method of production based on the original method of Peloquin and Hougas. However, dihaploids are usually male-sterile, unlike two of the first four, US-W1 and US-W4 [28][29][29,30], and female fertility can also be a problem in breeding [27][28]. Most dihaploids, like the cultivated diploid groups, display gametophytic self-incompatibility [30][31][31,32], which prevents the production of true breeding lines by self-pollination. In other words, diploid potatoes are outbreeders, and the same is true for most of their diploid wild relatives. The self-incompatibility (S) locus was mapped to chromosome 1 by Gebhardt et al. [32][33] and Kaufmann et al. [33][34]. In summary, there is no shortage of diploid germplasm available for diploid F1 hybrid breeding, but, as in all breeding programs, it will be important to use the best germplasm available for a particular target environment and particular end use. Breeding objectives will no doubt be expressed in terms of improving yield potential and tolerance to important abiotic stresses, and improving quality traits and resistance to important pests and diseases.4. Breeding Methods for Diploid Potatoes
Breeding diploid potatoes for clonal propagation can be conducted through the testing, selection, and vegetative multiplication of desirable clones from population improvement schemes, such as mass or family selection [16][17][18][19][20][34][17,18,19,20,21,35], or from pair crosses between clones that complement each other for desirable traits [22][23]. The new cultivar will have come from combining a gamete from its female parent with one from its male parent. For each of the 12 pairs of chromosomes, the female-derived chromosome will need to complement the male-derived chromosome for genetic loci at which they have different alleles. In the following simple example, small letters represent deleterious recessive alleles (Figure 1).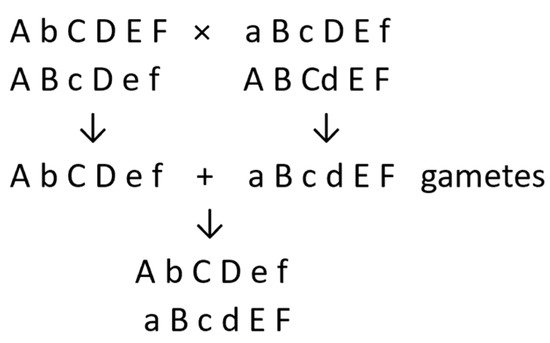
Figure 1.
New cultivar from cross between two heterozygous diploid parents (one pair of chromosomes shown, where small letters represent deleterious recessive alleles).
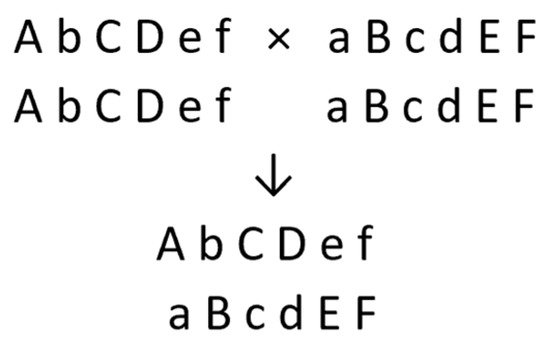
Figure 2. New diploid F1 hybrid cultivar from cross between two homozygous diploid parents (one pair of chromosomes shown, where small letters represent deleterious recessive alleles).
Despite being homozygous for some deleterious alleles, the inbred parents must have sufficient vigor and fertility for the maintenance and production of their hybrid.
The prerequisites for successful F1 hybrid breeding in plants are: (1) the ability to produce seed by self-pollination; (2) the ability to produce homozygous inbred lines with acceptable vigor and fertility, or sufficiently homozygous inbred lines to produce an F1 hybrid of acceptable phenotypic uniformity; (3) the ability to produce sufficient inbred lines for combinations to be found that are superior to existing cultivars, and to achieve this over cycles of inbreeding and crossbreeding for continued progress; and (4) the ability to produce large quantities of F1 seed for growing the hybrids commercially.