Epilepsy is a chronic disease that affects millions of people worldwide. Antiepileptic drugs (AEDs) are used to control seizures. Even though parts of their mechanisms of action are known, there are still components that need to be studied. Therefore, the search for novel drugs, new molecular targets, and a better understanding of the mechanisms of action of existing drugs is still crucial. Levetiracetam (LEV) is an AED that has been shown to be effective in seizure control and is well-tolerable, with a novel mechanism of action through an interaction with the synaptic vesicle protein 2A (SV2A). Moreover, LEV has other molecular targets that involve calcium homeostasis, the GABAergic system, and AMPA receptors among others, that might be integrated into a single mechanism of action that could explain the antiepileptogenic, anti-inflammatory, neuroprotective, and antioxidant properties of LEV. This puts it as a possible multitarget drug with clinical applications other than for epilepsy.
- levetiracetam
- antiepileptic drugs
- SV2A
- neuroprotection
- neuroinflammation
- GABAergic system
- calcium homeostasis
1. Introduction
2. Levetiracetam Binding Site (LBS)
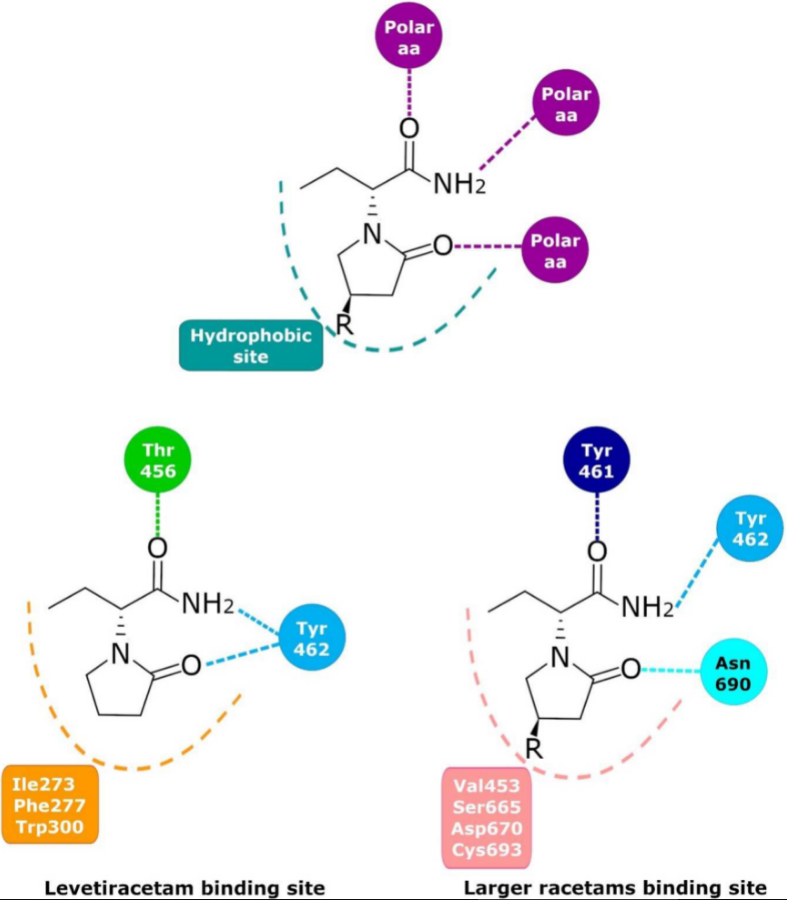
Since 1995, the presence of a specific LBS that could be involved in the anticonvulsant properties of LEV has been investigated; photoaffinity labeling studies determined that UCB-30889 (the LEV analog with a 30-fold greater affinity for SV2A) was bound to a protein of ~90 kDa. This protein was only detected in brain structures but not in peripheral tissue and was abundant in synaptic plasma membranes and in the synaptic vesicle fractions [99,100,101][18][19][20]. This placed SV2 proteins as the primary candidates [102][21]. Several studies demonstrated that UCB 30889 did not bind to brain membranes and purified SVs from mice lacking SV2A, indicating that SV2A is necessary for LEV binding. Moreover, UCB 30889 bound to SV2A but not to SV2B or SV2C proteins expressed in fibroblasts, indicating that SV2A is sufficient for LEV binding [9]. Currently, there is no doubt that LEV binds in a saturable, reversible, and stereospecific manner to SV2A in both rat and human brains; this is also true for its structural analogs [9,103][9][22]. Mutagenesis studies, molecular models, and molecular dynamics simulations have allowed important SV2A residues relevant for the LBS and its intermolecular interactions to be identified. Shi et al. found 14 residues that, when mutated, alter the binding of LEV to SV2A, they were: F277A, W300A, W300F, Y462A, K694A, G303A, F658A, V661A, I663A, W666A, N667A, S294A, M301A, and G659A [104][23]. Later, Lee et al. using two conformational states of the in silico model of SV2A, observed two additional residues, W454 and D670, that may contribute to LBS [105][24]. Finally, Correa-Basurto et al. identified additional hydrophobic and hydrogen bond interactions with T456, S665, and L689 (besides W454 and D670), which may be important for ligand recognition within the LSB and support the putative LBS observed previously [106][25] where the residues found by single aminoacid mutants [104][23] are distributed in a large volume which comprises the sites found by molecular modeling [105,106][24][25] (Figure 21). Moreover, by means of radioligand binding assays, it has been suggested that LEV and BRIV could have a different binding site or favor different conformational states of the SV2A protein [107][26], with the subsequent identification of the outward SV2A conformation relevant for differential binding of LEV (Ile273, Lys694, and Ser294) and BRIV (Lys694) [108][27]. Moreover, studies with the UCB-1244283 (SV2A positive allosteric modulator) indicated that the SV2A protein contains multiple interacting binding sites [91][28]. Thus, there is still a wide field of opportunities for the study of the molecular behavior of SV2A–ligand interactions (Figure 21).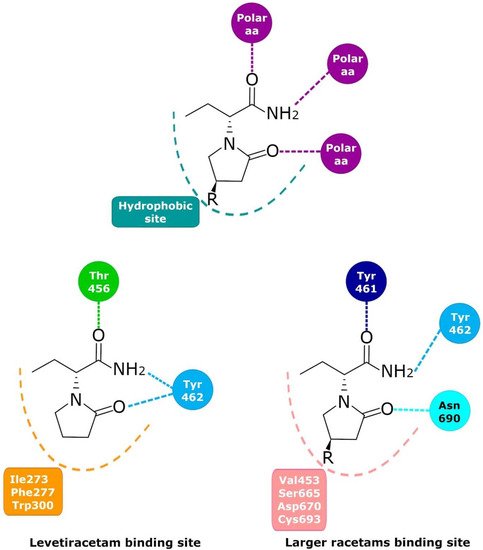
3. Molecular Mechanism
As mentioned above, different lines of evidence suggest that SV2A modulation constitutes the primary mechanism of action of LEV. However, other targets, for example, Alpha-Amino-3-Hidroxy-5-Methyl-4-Isoxazole Propionic Acid (AMPA), noradrenaline, adenosine, and serotonin receptors, or those involved in calcium homeostasis, the gamma-aminobutyric acid (GABA) system, and intracellular pH regulation may contribute to the effects of LEV.3.1. Synaptic Vesicle Protein 2A (SV2A)
SV2A is an integral membrane protein found in the vesicles of almost all synaptic terminals. In the synaptic vesicle cycle, several functions in both exocytosis and endocytosis processes have been attributed to SV2A. During exocytosis, the SV2A protein might function first as a target for residual Ca2+ (Figure 32); by means of paired pulses and the repetitive stimulation of 2 and 10 Hz in SV2A/SV2B double knockout (DKO) neurons, an increase was observed in the synaptic response (relative facilitation) with subsequent progressive depression. Such a facilitation was reversed in all frequencies by incubating DKO neurons with slow calcium buffer egtazic acid (EGTA), indicating that the difference in synaptic response was mostly due to the accumulation of residual Ca2+ [109,110,111][29][30][31]. Second, as a facilitator of the progression to the release-competent state in the vesicular priming (Figure 32). SV2A knockout (KO) or SV2A/SV2B DKO hippocampal neurons, as well as knockdown presynaptic SV2A in superior cervical ganglion neurons, resulted in a decrease in neurotransmitter release and a reduced size of the readily releasable pool (RRP) of vesicles, suggesting that SV2A maintains normal neurotransmission by regulating the RRP size [112,113,114][32][33][34] (Figure 32). In addition, brain tissue from SV2A KO contained a smaller proportion of the syntaxin protein, consistent with the interpretation that the loss of SV2A affects the formation of the complex of soluble NSF attachment proteins receptor (SNARE; [113,115][33][35]. Since the absence of SV2A presented a normal number of vesicles docked at the active zone in presynaptic terminal [113,115][33][35] and the mechanism of exocytosis itself was not affected (no changes were observed in any SV2A KO and SV2A/SV2B DKO hippocampal neurons when examining miniature postsynaptic currents) [109[29][32][33][36],112,113,116], but there were diminished RRP and the formation of the SNARE complexes was affected, SV2A may influence the synaptic vesicle priming step in the synaptic vesicle cycle, maintaining the availability of secretory vesicles and thus the release probability; thereby, ensuring correct neurotransmission. Third, during endocytosis, SV2A may regulate the vesicle content of the calcium-sensor synaptotagmin (SYT-1; Figure 32). Mutation in Y46 amino acid residue (an endocytosis motif) at the NH2 -terminus of SV2A (where SYT-1 binds to SV2A) caused a higher proportion of both SV2A and SYT-1 on the plasma membrane, indicating a reduced internalization. The Y46 residue of SV2A serves as a receptor for clathrin adaptor proteins; thus, SV2A via an interaction with both SYT-1 and clathrin adaptors may regulate the adequate trafficking of SYT-1 and, in consequence, the Ca2+-stimulated fusion [111,117,118][31][37][38].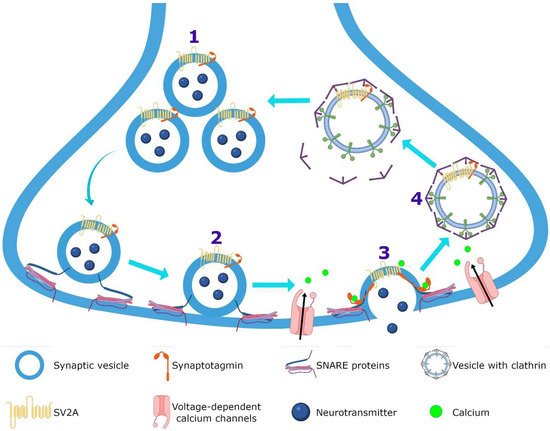
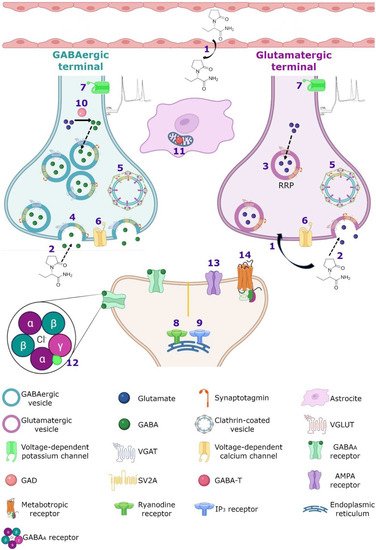
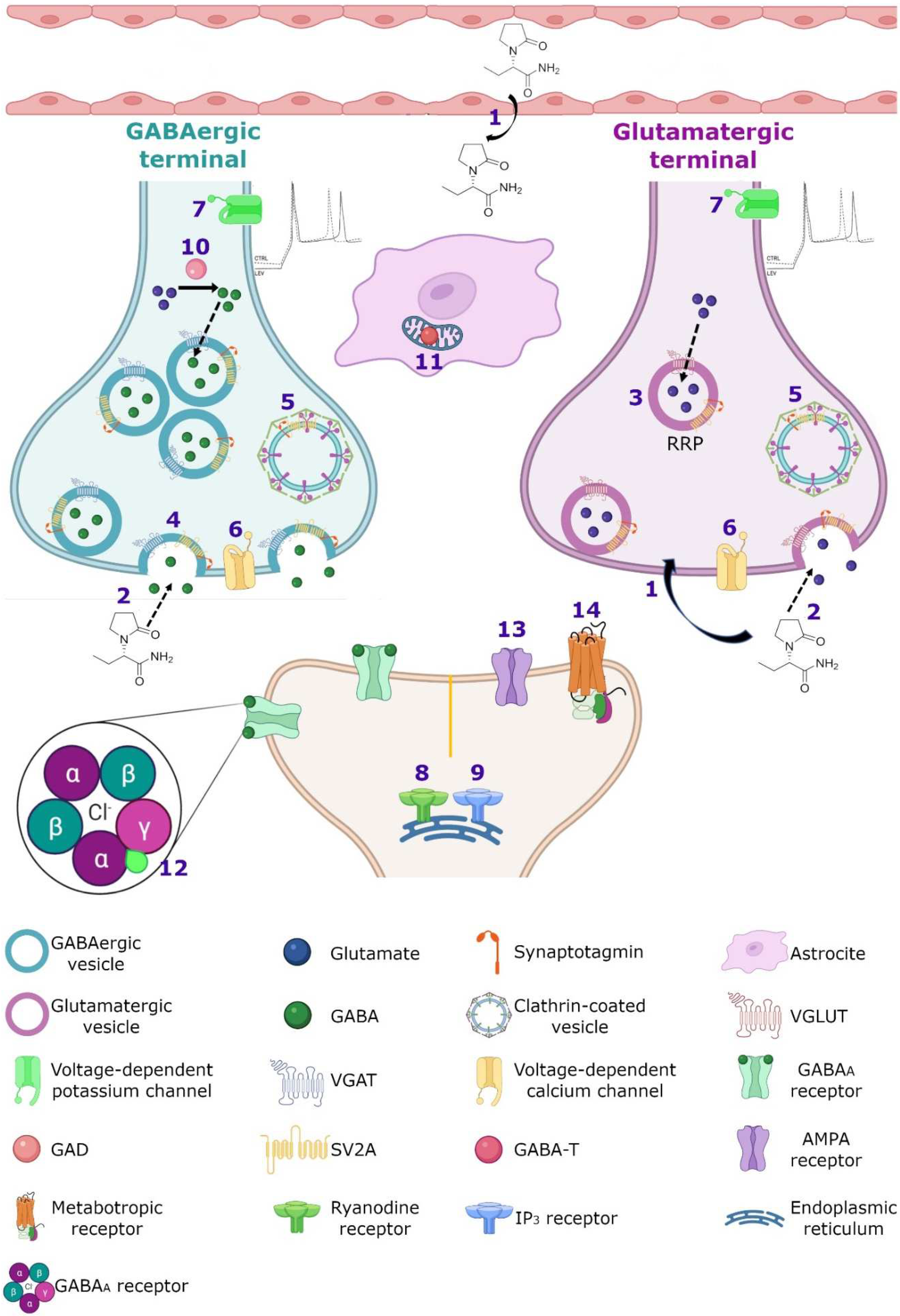
3.2. Calcium Homeostasis
There is significant experimental evidence showing that LEV modulates targets related with cellular Ca2+ which is a ubiquitous signal transduction molecule that plays a key role in the modulation of neuronal excitability and synaptic transmission. Specifically, LEV effects have been observed in voltage-gated channels and Ca2+ signaling. Several studies have shown that LEV can block the high-voltage activated (HVA) Ca2+ channels N-type, P/Q-type and L-type (Figure 43). The administration of LEV (32 µM) in CA1 of rat hippocampal slices decreased significantly the neuronal HVA Ca2+ currents [135][55]. Other studies showed a selective LEV inhibition of N-type Ca2+ channels in isolated striatal, neocortical, and CA1 pyramidal hippocampal neurons [136,137,138][56][57][58]. In addition, LEV (100 µM) provoked a partial reduction in P/Q-type HVA Ca2+ currents in the acutely isolated neocortical neurons [137][57]. Moreover, LEV inhibited Ca2+ entry by blocking the type Ca2+ L-type channels in hippocampal CA3 neurons obtained from spontaneously epileptic rats [139][59]. This effect was more potent than that in control neurons, suggesting that this may contribute to the antiepileptic effect of LEV [139,140][59][60]. Moreover, it has been reported that LEV elicits effects on HVA Ca2+ channels (presumably N-type) of superior cervical ganglion cholinergic neurons. Data showed that LEV inhibited synaptic transmission between these cells in a time-dependent manner, significantly reducing excitatory postsynaptic potential (EPSP) after a 1 h of application. Interestingly, intracellular LEV administration caused (after 4 to 5 min of exposition) rapid inhibition of the Ca2+ current; this is consistent with a mechanism where LEV may interact directly with HVA Ca2+ channels, causing a reduction in synaptic transmission [19,114][34][61]. Furthermore, the application of LEV (100 μM) in acutely isolated hippocampal CA1 neurons from rats and guinea pigs, reduced the delayer rectifier K+ currents by 26%, causing a decrease in the repetitive action potential generation and subsequent, leading to a slight prolongation of duration of the first action potential. Thus, LEV action may also be also related to its ability to hyperpolarize the membrane potential via K+ channel activation [11] (Figure 43). On the other hand, various studies have reported that LEV is an effective inhibitor of Ca2+ release mediated by the two of the major systems of calcium-induced calcium release, ryanodine and inositol-3-phosphate (IP3) receptors (Figure 43). LEV significantly reduced the Ca2+ transients induced by caffeine (a ryanodine receptor activator) in cultured rat hippocampal neurons [141,142][62][63]. In addition, LEV inhibited the epileptiform effect induced by caffeine on the evoked field potentials and delayed caffeine-induced spontaneous bursting on rat hippocampal slices [141][62]. Moreover, LEV inhibited Ca2+ transients induced by bradykinin (BK; a stimulator of IP3 receptor) in hippocampal neurons, causing a 74% diminution in calcium release mediated by the IP3 receptor compared to the control [142][63]. In PC12 rat pheochromocytoma cells, LEV decreased in a dose-dependent manner, the increase in Ca2+ caused by the application of 1µM of bradykinin or 100 µM of ATP. The inhibitory effect of LEV was mainly exerted by IP3-triggered Ca2+ store depletion without reducing Ca2+ storage into these deposits [143][64]. Then, the ability of LEV to modulate ryanodine and IP3 receptors demonstrated another important molecular effect of this agent on a major second messenger system in neurons.3.3. GABAergic System
The GABA is the main inhibitory neurotransmitter of the central nervous system. Multiple AEDs act on its GABAA receptor to increase inhibition and thereby controlling the aberrant neuronal activity and seizures [144][65]. Regarding LEV, however, there are conflicting results concerning its effect on the GABAergic system [145][66]. Patients with two distinct pathologies, focal epilepsy or migraine, were treated with LEV to determine if this drug modified brain GABA levels. By means of proton magnetic resonance spectroscopy, the GABA/creatinine ratio was evaluated before and during treatment of patients with epilepsy. The data showed an increase in GABA/creatinine in the occipital lobe in responder patients (those who showed 50–100% seizure reduction), while in non-responders the results were inconclusive [146][67]. Meanwhile, migraine patients treated with LEV showed a decrease in headache frequency and intensity associated with a decrease in posterior cingulate cortex GABA levels [147][68]. These results suggest that LEV may modulate differentially GABA levels and these results agreed with several animal studies. For example, the administration of LEV before injecting the convulsive agent pilocarpine in a murine model, protected against seizures modulating different neurotransmitter release; hence, LEV reversed alterations induced by focal to bilateral tonic-clonic seizures, increasing aspartate and reducing glutamine, GABA, and glycine levels in rat hippocampus [148][69]. In addition, by K+-evoked depolarization with microdialysis technique, LEV inhibited the release of biogenic amines, GABA, and L-glutamate in the medial prefrontal cortex of control rats [149][70]; however, in epileptic rats treated with LEV for one week, the K+-evoked depolarization induced a preferential increase in GABA levels without modifying other neurotransmitters in the rat dorsal hippocampus [150][71]. In addition, the administration of LEV in the substantia nigra, a mainly GABAergic nucleus, showed that this anticonvulsive drug decreased the spontaneous firing of non-dopaminergic (maybe GABAergic) neurons, suggesting that the modulation of neuronal firing in GABAergic projections from the substantia nigra, could involve the activation or inhibition of neurotransmitter systems in other brain areas [148,151][69][72]. In temporal lobe epilepsy (TLE) both in animal models and patients, a run-down current elicited by GABA, which is disease progression-dependent, has been reported [152,153][73][74]; the repetitive activation of GABAA receptors induces a decrease in GABAergic signaling (current) use-dependent in hippocampal and cortical neurons denominated as run-down, this desensitization of GABAA receptor, could increase hyperexcitability and favor the occurrence of seizures [152,153][73][74]. In oocytes microtransplanted with ionotropic GABAA-receptors obtained from the resected hippocampus and temporal neocortex of patients with mesial TLE, as well as rats with pilocarpine-induced TLE, the run-down of the current evoked by GABA and the effect of LEV on this current were assessed [153,154][74][75]. In chronic epilepsy, both rats and patients showed an increase in current GABA run-down in the hippocampus and cortex and LEV had a region-dependent effect [153,154][74][75]; in the tissue of these rats, incubation with LEV did not affect the run-down current in the hippocampus, but it does attenuate it in the cortex [153][74]. Meanwhile, in the tissue of patients, LEV inhibited the GABA-current run-down in the hippocampus and neocortex but was ineffective in the hippocampal subiculum [154][75]. The authors argue that the differences could be because in the subiculum of mesial TLE patients, a switch is generated in GABA where it becomes an excitatory neurotransmitter, while in the hippocampus and neocortex GABA functions as the classic inhibitory neurotransmitter, another option to explain this data, is the differential subunit composition of GABAA receptors and their phosphorylation [154][75]. However, there are controversial results regarding the effect of LEV in the metabolism of GABA, since on in vitro assays LEV did not alter the activity of the GABA synthesizing enzyme, glutamic acid decarboxylase (GAD) or the GABA degrading enzyme and GABA-transaminase (GABA-T); however, on in vivo studies, a decrease in GAD in striatum and an increase in the hypothalamus at high doses of the drug was observed, as well as an increase in GABA-T activity in the cortex, striatum, thalamus and cerebellum [151][72] (Figure 43). Moreover, LEV modified GABA turnover by reducing it in the striatum but increasing it in the cortex and hippocampus [151][72]. A reduction in GAD and GABA turnover in the striatum might disinhibit GABAergic striatal output pathways, augmenting the inhibition in their respective target regions and increasing the anticonvulsant effects of LEV [151][72]. However, the authors postulate that those differential alterations on GAD and GABA-T activity between both models and in different regions might not be directly caused by LEV, instead as a consequence to pre or postsynaptic secondary effects [151][72]. Another action of LEV in the GABAergic system involves the ability to reverse the inhibitory effects of the negative allosteric modulators β-carbolines and zinc on both GABAA and glycine receptors, the two main ionotropic inhibitory receptor systems in the brain [145][66]. β-carbolines congeners can act at the benzodiazepine recognition site of the GABAA receptor complex to inhibit GABA-stimulated chloride conductance (inverse agonist effect) [155,156][76][77] (Figure 43). By using the whole cell patch-clamp technique, it was observed that LEV reversed the inhibitory effect of the methyl-6,7-dimethoxy-4-ethyl-β-carboline-3-carboxylate (DMCM, inverse agonist), N-methyl-beta-carboline-3-carboxamide (FG7142, partial Inverse agonist), and butyl 9H-pyrido[3,4-b]indole-3-carboxylate (β-CCB, inverse agonist) on GABA-elicited currents in hippocampal and cerebellar granule neurons [145][66]. In addition, LEV completely abolished the inhibitory effects of DMCM and β-CCB on glycine currents of spinal neurons [145][66]. The in vitro interaction of LEV with negative allosteric modulators of inhibitory receptors was confirmed in vivo in sound-susceptible mice; the administration of LEV (17 mg/kg) produced an important suppression of convulsions in these mice. The protective LEV effect was significantly diminished by the co-administration of FG 7142, from a dose of 5 mg/kg [145][66]. In addition, in postnatal day 10 rat pups, treatment with LEV decreased the severity of DMCM-evoked seizures in a dose-dependent manner when administered in doses of 10 mg/kg and greater [157][78]. Moreover, LEV completely reversed the inhibition by zinc of GABA and glycine evoked currents, in hippocampal and spinal cord neurons [145][66]. By recording functional synaptic-boutons, it was observed that the activation of GABAA receptors by muscimol (a selective GABAA receptor agonist) induced the inhibition of evoked excitatory postsynaptic currents (eEPSCs); later, in the continued presence of muscimol, the addition of Zn2+ increased the eEPSC amplitude (Zn2+ had no effect by itself on the eEPSC). However, when LEV was applied in the continuous presence of muscimol and Zn2+, there was a decrease in the eEPSC amplitude (also, LEV had no effect by itself on eEPSC, and in the presence of muscimol without Zn2+). Then, LEV reversed the Zn2+ induced suppression of GABAA receptors, resulting in a decrease in glutamatergic excitatory transmission [158][79]. These results suggest that, also, the antagonism of allosteric Zn2+ modulation by LEV may be one of its mechanisms of action.3.4. SV2A and GABAergic System
Despite of the fact that the SV2A protein is localized in all synaptic vesicles regardless of neurotransmitter content and its expression is similar on both glutamatergic and GABAergic terminals [159[80][81][82][83],160,161,162], a strong relationship between SV2A and the GABAergic system has been observed. In a microdialysis study on rats with a mutation of the SV2A gene (SV2AL174Q), a decrease in depolarization-evoked GABA release in hippocampus and amygdala was shown, without modification of the levels of glutamate [163,164][84][85]. In addition, in a rat model of SE induced by pilocarpine, an increase in hippocampal SV2A expression associated with GABAergic but no with glutamatergic terminals was reported [165][86]. Moreover, recordings of cultured hippocampal pyramidal neurons (CA1 and CA3) from SV2A KO and SV2A/SV2B DKO mice showed a decreased frequency and amplitude of spontaneous inhibitory postsynaptic currents (sIPSCs) and an increase in the frequency of the spontaneous excitatory postsynaptic currents (sEPSCs) but without any change in their amplitude [112,116][32][36]. These data agree with the observations of a great co-expression between SV2A and GABAergic neurons in the amygdala and hippocampus [163,164,166,167][84][85][87][88]. Finally, systemic administration of LEV decreased the hyperalgesia, probably enhancing GABAergic neurotransmission by different pathways, but the local administration of LEV did not act on the GABA system [168,169][89][90]. The close association between SV2A and the GABAergic system could be very important for the effects of LEV, but further studies must be completed in order to clarify this issue.3.5. AMPA Receptors
Regarding the effect of LEV on glutamatergic receptors, few studies have been performed; in recordings of neuronal cortical cultures by whole cell patch-clamp, the application of kainate induced inward currents in all neurons, which were mediated primarily by the activation of AMPA receptors. The incubation with LEV in this culture, decreased kainate-induced currents by 26.5% and returned to basal after LEV washout, indicating a mild modulation of AMPA receptors [170][91]. This was confirmed by the administration of cyclothiazide, an AMPA positive allosteric modulator in these cells, since LEV decreased the amplitude of currents induced by cyclothiazide as well as diminished the amplitude and frequency of mEPSC [170][91].3.6. Noradrenaline, Adenosine and Serotonin Receptors
LEV has also shown direct or indirect interaction with noradrenaline (α2A- and α2C), adenosine (A1), and serotonin (5-HT1B/1D) receptors contributing to its anti-hyperalgesic effect [11]. In an intraplantar carrageenan-induced model of inflammatory pain, the antagonists CTAP (µ-opioid receptor antagonist), BRL-44408 (α2A-adrenoceptor antagonist), MK-912 (α2C-adrenoceptor antagonist), 1,-3-dypropyl-8-cyclopentylxanthine, DPCPX (adenosine A1 receptor antagonist), GR-127935 (5-HT1B/1D receptor antagonist) and bicuculline (GABAA receptor antagonist) were injected (i.p. or intraplantarly) before LEV administration (systemic, 10–200 mg/kg or local, 200–1000 nmol/paw); subsequently, it was assessed if LEV had an effect by means of paw pressure test, over time (60–300 min) [168,169][89][90]. The authors reported that LEV exerted dose and time-dependent anti-hyperalgesic activity. In contrast, the administration of all antagonists decreased this effect of LEV [168,169][89][90]. These results suggest that LEV could be a promising drug for inflammatory pain in humans.3.7. Intracellular pH Regulation
Variations in the normal intracellular pH influence diverse functions in neurons, glia, and interstitial space [171][92]. Intracellular pH regulation is grouped into acid extrusion and acid loading. Acid extrusion is mainly accomplished by Na+/H+ exchangers, Na+-dependent Cl−/HCO3− (chloride/bicarbonate) exchangers, and Na+/HCO3− (sodium/bicarbonate) co-transporters. Acid loading is mediated by Na+-independent Cl−/HCO3− exchangers [171,172][92][93]. Several studies have suggested that the Na+-independent Cl−/HCO3− exchanger, AE3, may modulate seizure susceptibility [171,173][92][94]. Moreover, it has been proposed that LEV mediates pH shifts and seizure-like activity via HCO3− regulation. In human neocortical brain slices from patients with TLE pharmaco-resistant, LEV was associated with a subtle acidification, predominantly in more alkaline cells. This acidification was depended on the extracellular HCO3− concentration [20][95]. Then, since acidifications induced by LEV were based upon an inhibition of the Na+/HCO3− co-transporters, Na+-dependent HCO3− transporters and Na+-dependent Cl−/HCO3− exchangers, LEV may decrease the intracellular pH by weakening the transmembrane HCO3−-mediated acid extrusion [20,174][95][96]. In addition, recordings from hippocampal slices treated with 4-aminopyridine (to increase neuronal excitability), showed that administration of LEV (10–100 µM), decreased the frequency of spontaneous action potentials and bursts of CA3 neurons. Both effects were reversible upon LEV washout and by incubating LEV plus the alkalinizing agent trimethylamine [174][96]. Then, by inducing intracellular acidification, LEV may attenuate the excitatory neuronal activity, promoting the termination of epileptic activity and contributing to its anticonvulsive potency [20][95].3.8. Single or Integrated LEV Molecular Mechanism of Action?
Traditionally, the SV2A protein has been considered as the main therapeutic target of LEV. Löscher et al. proposed that although some cellular and molecular effects of LEV may contribute to its unique pharmacological profile, they have a modest magnitude [8]. However, Cortes et al. pointed out that the mechanism of action of LEV comprises a cascade of effects that in the first instance, are exerted by binding to the SV2A protein, but its pharmacodynamics involve various molecular targets that must be integrated into a single mechanism of action by a single pathway [11]. WResearchers agree with this last point of view and further propose that LEV is a unique antiepileptic agent that has multiple mechanisms of action that from an integrated point of view may explain not only its molecular effects but also their genetic, antiepileptic, antiepileptogenic, neuroprotective, antioxidant and anti-inflammatory actions (see below). Figure 43 represents the hypothetical integrated molecular mechanisms of action of LEV.4. Genetic Mechanism
The antiepileptic activity of LEV has been related, besides the SV2A expression, to modifications in the expression of diverse genes. In amygdala-kindled rats, this process was associated with an upregulation of hippocampal brain-derived neurotrophic factor (BDNF) and neuropeptide Y (NPY) mRNA levels. Treatment for 12 days with LEV clearly delayed the progression of kindling, showing a clear antiseizure effect and prevented the increase in BDNF and NPY mRNA [175,176][97][98]. In addition, using real-time quantitative polymerase chain reaction temporal lobe expression of NPY gene and other epilepsy-related genes, such as, thyrotropin-releasing hormone (TRH) and glial fibrillary acidic protein (GFAP) were confirmed to be up-regulated in amygdala-kindled rats and partially normalized by LEV treatment [175][97]. In another work using amygdala-kindled rats, LEV 1 h prior to the kindling stimulation attenuated the hippocampal overexpression of TNF-α and Cox-2, two genes related to inflammatory processes. The decrease in the expression of both genes was parallel to the antiseizure effect of the drug [177][99]. LEV every day in 1 week also reduced the expression levels of interleukin-1β (IL-1β) and interleukin-1 receptor subtype I, and the associated reactive gliosis in the hippocampus and piriform cortex of epileptic rats [178][100]. In relation of the effect of LEV with other genetic pathologic mechanisms, Rassu et al. showed that LEV treatment ameliorated the effect of pathological mutant phenotype of leucine-rich repeat kinase 2 (LRRK2), an enzyme that controls the vesicle trafficking [179][101]. In thise study, LEV treatment significantly decreased the neurite shortening phenotype of mutant mice in primary neurons and in PC12 cells. LEV also diminishes the accumulation of dopamine receptor D2 (DRD2) into the Golgi areas due to the mutant expression in SH-SY5Y cells. These results indicated that LEV reverts LRRK2 G2019S-associated pathological effects and that LRRK2 and SV2A are involved in a common protein network controlled by LEV with the consequent modulation of traffic and dynamics in neurons [179][101]. In another study, it was showed that LEV once daily for 5 days reduced the effect of presynaptic gene Stxbp1 mutations in Stxbp1+/− mice, significantly reducing the number of spike-wave discharges. The de novo heterozygous mutations in STXBP1/Munc18-1 gene were implicated in the development of early infantile epileptic encephalopathies [180][102]. In a clinic case, LEV administered continuously from one month of age to 26 months of age had a dramatic efficacy in the treatment of encephalopathy refractory in a child with de novo heterozygous mutation (c.[922A>T]p.[Lys308(∗)]) in the STXBP1 gene [181][103]. Moreover, it was shown that the administration of LEV (mean duration of 50.59 ± 37.93 months) in women with a diagnosis of epilepsy caused high serum levels of the Wnt antagonists sclerostin and DKK-1 in comparison with the healthy controls. Thise study showed that the LEV effect is implicated the modulation of the Wnt signaling pathway [182][104]. In addition, it has been shown that LEV (18 and 180 μM) significantly decreases the gene expression of excitatory amino acid transporter 2, (EAAT2) in brain metastasis glioblastoma cells culture, indicating that LEV has a mechanism of action that decreases the recapture of glutamate from the extracellular space in brain cancer [183][105]. In addition, in a mouse microglial BV-2 cell line culture, it was shown that lipopolysaccharide increases activator protein-1 (AP-1), FOS like 1, AP-1 transcription factor subunit (FosL1), MAF BZIP transcription factor F (MAFF) and Spi-C transcription factor (SPIC) mRNA levels and LEV attenuated AP-1 and FosL1 mRNA expression in this model. Therefore, the authors suggested that LEV can be a candidate for the treatment of neurological diseases that involve microglial activation [184][106]. Neuronal activity influences gene expression and the drugs that modify it, through interference in neurotransmission. Therefore, LEV can affect gene expression [122,185,186][42][107][108]. There is evidence that in HeLa cells, the main metabolite of LEV blocks histone deacetylases which catalyze the hydrolysis of acetyl groups from the lysine of some proteins, such as histone tails, inducing chromatin condensation and inhibiting gene transcription [187][109]. In addition, it was observed in electrocorticography registers of epileptic patients an epileptiform electrical activity in the form of spikes, which has been associated with changes in gene expression [188,189][110][111]. Thus, if LEV can inhibit hypersynchronous neuronal activity by reducing the epileptiform activity-induced population spikes in CA3 [148][69] and in dentate gyrus (DG) [190][112], this suggests that LEV could modulate the expression of genes, as wresearchers mentioned above.Effect of Gene Polymorphisms in LEV Treatment in Clinical Studies
Several investigators have studied the association of some gene polymorphisms with LEV treatment (with a target dose of 20–60 mg·kg−1 daily for 3–4 weeks) in epileptic population. Zhao et al. studied the impact of adenosine-triphosphate (ATP)-binding cassette sub-family B member 1 (ABCB1) polymorphisms rs1128503 (C1236T), rs2032582 (G2677T) and rs1045642 (C3425T) in exon 26 on LEV serum levels and treatment efficacy in 245 Uygur Chinese children with epilepsy (drug resistant and drug-responsive). The authors showed that for genotype frequencies of ABCB1 G2677T/A, the GT genotype frequency was significantly different between drug-resistant and drug-responsive groups (OR = 0.484, 95%CI = 0.236–0.003, p = 0.046). The other genotype frequencies did not significantly differ between both groups [191][113]. In relation to the serum drug concentration of LEV and the serum drug concentration/body mass dose ratios (CDR), it was observed that rs2032582 and rs1045642 polymorphisms are significantly related to LEV concentration and CDR values. A higher LEV concentration and CDR values were found in GT, TT, GA, and AT genotype carriers of rs2032582 polymorphism in comparison with GG carriers (p = 0.021). Higher LEV concentrations and CDR values were found in TT genotype carriers of rs1045642 compared with CC and CT carriers (p = 0.002). The authors concluded that ABCB1 rs2032582 and rs1045642 polymorphisms may affect the therapeutic efficacy of LEV in epilepsy [191][113]. In relation to SV2A mutations, surprisingly, in a pediatric epileptic patient with a rare de novo heterozygous variant in SV2A (NM_014849.5:c.1978G>A:p.Gly660Arg), seizures were found to worsen after treatment with increasing doses of LEV for approximately 40 days [192][114]. Moreover, Wolking et al. investigated the genetic risk of rare variants for drug response to three AEDs (including LEV treatment for at least 1 year) through an analysis of the sequencing, genotyping, variant selection, and annotation of an epileptic cohort derived from the EpiPGX Consortium, to identify genetic biomarkers of epilepsy treatment response and adverse drug reactions [193][115]. The individuals who met the inclusion criteria were 1622 patients with a diagnosis of focal epilepsy and genetic generalized epilepsy. The results of the gene set analysis showed a significant enrichment of protein truncating variants and splice-region variants in the SV2 gene group (SV2A and SV2B) associated with drug resistance. Thereafter, it was possible to conclude that a group of genes are related to drug kinetics or targeting in drug resistance to LEV [193][115]. In another study, through the screening for mRNA signatures in 53 epileptic hippocampal tissue from pharmacoresistant TLE patients, abundant synapse-associated molecule mRNA signatures in LEV a priori non-responders were shown. In the promoter characterization was observed an accumulation of the rs9305614 G-allele in phosphatidylinositol N-acetylglucosaminyltransferase promoter with activation of LBP-1 transcription factor of LEV a priori non-responders mesial TLE patients. The authors suggest that epigenetic factors predisposing for a priori LEV pharmacoresistance by transcriptional targets [194][116]. Finally, in another study, the presence of rs1611115, rs4680 and rs1800497 polymorphisms with adverse psychotropic side effects of LEV in patients with chronic epilepsy was found to be related with a decrease in dopaminergic activity [195][117].References
- World Health Organization (WHO). Epilepsy. Available online: https://www.who.int/news-room/fact-sheets/detail/epilepsy (accessed on 6 October 2021).
- Fisher, R.S.; Acevedo, C.; Arzimanoglou, A.; Bogacz, A.; Cross, J.H.; Elger, C.E.; Engel, J.J.; Forsgren, L.; French, J.A.; Glynn, M.; et al. ILAE official report: A practical clinical definition of epilepsy. Epilepsia 2014, 55, 475–482.
- Fisher, R.S.; Cross, J.H.; French, J.A.; Higurashi, N.; Hirsch, E.; Jansen, F.E.; Lagae, L.; Moshé, S.L.; Peltola, J.; Roulet Perez, E.; et al. Operational classification of seizure types by the International League Against Epilepsy: Position Paper of the ILAE Commission for Classification and Terminology. Epilepsia 2017, 58, 522–530.
- Pérez-Pérez, D.; Frías-Soria, C.L.; Rocha, L. Drug-resistant epilepsy: From multiple hypotheses to an integral explanation using preclinical resources. Epilepsy Behav. 2021, 121, 106430.
- Pitkänen, A.; Sutula, T.P. Is epilepsy a progressive disorder? Prospects for new therapeutic approaches in temporal-lobe epilepsy. Lancet Neurol. 2002, 1, 173–181.
- Klein, P.; Friedman, A.; Hameed, M.Q.; Kaminski, R.M.; Bar-Klein, G.; Klitgaard, H.; Koepp, M.; Jozwiak, S.; Prince, D.A.; Rotenberg, A.; et al. Repurposed molecules for antiepileptogenesis: Missing an opportunity to prevent epilepsy? Epilepsia 2020, 61, 359–386.
- Alrabiah, H. Levetiracetam. Profiles Drug Subst. Excip. Relat. Methodol. 2019, 44, 167–204.
- Löscher, W.; Gillard, M.; Sands, Z.A.; Kaminski, R.M.; Klitgaard, H. Synaptic Vesicle Glycoprotein 2A Ligands in the Treatment of Epilepsy and Beyond. CNS Drugs 2016, 30, 1055–1077.
- Lynch, B.A.; Lambeng, N.; Nocka, K.; Kensel-Hammes, P.; Bajjalieh, S.M.; Matagne, A.; Fuks, B. The synaptic vesicle is the protein SV2A is the binding site for the antiepileptic drug levetiracetam. Proc. Natl. Acad. Sci. USA 2004, 101, 9861–9866.
- Crepeau, A.Z.; Treiman, D.M. Levetiracetam: A comprehensive review. Expert Rev. Neurother. 2010, 10, 159–171.
- Cortes-Altamirano, J.L.; Olmos-Hernández, A.; Bonilla-Jaime, H.; Bandala, C.; González-Maciel, A.; Alfaro-Rodríguez, A. Levetiracetam as an antiepileptic, neuroprotective, and hyperalgesic drug. Neurol. India 2016, 64, 1266–1275.
- Wong, L.C.; Freeburg, J.D.; Montouris, G.D.; Hohler, A.D. Two patients with Hashimoto’s encephalopathy and uncontrolled diabetes successfully treated with levetiracetam. J. Neurol. Sci. 2015, 348, 251–252.
- Rossi, S.; Mataluni, G.; Codecà, C.; Fiore, S.; Buttari, F.; Musella, A.; Castelli, M.; Bernardi, G.; Centonze, D. Effects of levetiracetam on chronic pain in multiple sclerosis: Results of a pilot, randomized, placebo-controlled study. Eur. J. Neurol. 2009, 16, 360–366.
- Falah, M.; Madsen, C.; Holbech, J.V.; Sindrup, S.H. A randomized, placebo-controlled trial of levetiracetam in central pain in multiple sclerosis. Eur. J. Pain 2012, 16, 860–869.
- Steinhoff, B.J.; Staack, A.M. Levetiracetam and brivaracetam: A review of evidence from clinical trials and clinical experience. Ther. Adv. Neurol. Disord. 2019, 12, 3518.
- Kenda, B.M.; Matagne, A.C.; Talaga, P.E.; Pasau, P.M.; Differding, E.; Lallemand, B.I.; Frycia, A.M.; Moureau, F.G.; Klitgaard, H.V.; Gillard, M.R.; et al. Discovery of 4-substituted pyrrolidone butanamides as new agents with significant antiepileptic activity. J. Med. Chem. 2004, 47, 530–549.
- Leclercq, K.; Matagne, A.; Provins, L.; Klitgaard, H.; Kaminski, R.M. Pharmacological Profile of the Novel Antiepileptic Drug Candidate Padsevonil: Characterization in Rodent Seizure and Epilepsy Models. J. Pharmacol. Exp. Ther. 2020, 372, 11–20.
- Noyer, M.; Gillard, M.; Matagne, A.; Hénichart, J.P.; Wülfert, E. The novel antiepileptic drug levetiracetam (ucb L059) appears to act via a specific binding site in CNS membranes. Eur. J. Pharmacol. 1995, 286, 137–146.
- Gillard, M.; Fuks, B.; Michel, P.; Vertongen, P.; Massingham, R.; Chatelain, P. Binding characteristics of ucb 30889 to levetiracetam binding sites in rat brain. Eur. J. Pharmacol. 2003, 478, 1–9.
- Rogawski, M.A. Brivaracetam: A rational drug discovery success story. Br. J. Pharmacol. 2008, 154, 1555–1557.
- Bajjalieh, S.M.; Peterson, K.; Shinghal, R.; Scheller, R.H. SV2, a brain synaptic vesicle protein homologous to bacterial transporters. Science 1992, 257, 1271–1273.
- Gillard, M.; Fuks, B.; Leclercq, K.; Matagne, A. Binding characteristics of brivaracetam, a selective, high affinity SV2A ligand in rat, mouse and human brain: Relationship to anti-convulsant properties. Eur. J. Pharmacol. 2011, 664, 36–44.
- Shi, J.; Anderson, D.; Lynch, B.A.; Castaigne, J.-G.; Foerch, P.; Lebon, F. Combining modelling and mutagenesis studies of synaptic vesicle protein 2A to identify a series of residues involved in racetam binding. Biochem. Soc. Trans. 2011, 39, 1341–1347.
- Lee, J.; Daniels, V.; Sands, Z.A.; Lebon, F.; Shi, J.; Biggin, P.C. Exploring the interaction of SV2A with racetams using homology modelling, molecular dynamics and site-directed mutagenesis. PLoS ONE 2015, 10, e0116589.
- Correa-Basurto, J.; Cuevas-Hernández, R.I.; Phillips-Farfán, B.V.; Martínez-Archundia, M.; Romo-Mancillas, A.; Ramírez-Salinas, G.L.; Pérez-González, Ó.A.; Trujillo-Ferrara, J.; Mendoza-Torreblanca, J.G. Identification of the antiepileptic racetam binding site in the synaptic vesicle protein 2A by molecular dynamics and docking simulations. Front. Cell. Neurosci. 2015, 9, 125.
- Wood, M.D.; Gillard, M. Evidence for a differential interaction of brivaracetam and levetiracetam with the synaptic vesicle 2A protein. Epilepsia 2017, 58, 255–262.
- Wood, M.D.; Sands, Z.A.; Vandenplas, C.; Gillard, M. Further evidence for a differential interaction of brivaracetam and levetiracetam with the synaptic vesicle 2A protein. Epilepsia 2018, 59, e147–e151.
- Daniels, V.; Wood, M.; Leclercq, K.; Kaminski, R.M.; Gillard, M. Modulation of the conformational state of the SV2A protein by an allosteric mechanism as evidenced by ligand binding assays. Br. J. Pharmacol. 2013, 169, 1091–1101.
- Janz, R.; Goda, Y.; Geppert, M.; Missler, M.; Südhof, T.C. SV2A and SV2B function as redundant Ca2+ regulators in neurotransmitter release. Neuron 1999, 24, 1003–1016.
- Chang, W.-P.; Südhof, T.C. SV2 renders primed synaptic vesicles competent for Ca2+ -induced exocytosis. J. Neurosci. 2009, 29, 883–897.
- Pichardo, L.A.; Contreras, I.J.; Zamudio, S.R.; Mixcoha, E.; Mendoza, J.G. Synaptic Vesicle Protein 2A as a novel pharmacological target with broad potential for new antiepileptic drugs. In Antiepileptic Drug Discovery: Novel Approaches, Methods in Pharmacology and Toxicology, 1st ed.; Talevi, A., Rocha, L., Eds.; Springer: Berlin, Germany, 2016; pp. 53–65. ISBN 13:978-1493963539.
- Crowder, K.M.; Gunther, J.M.; Jones, T.A.; Hale, B.D.; Zhang, H.Z.; Peterson, M.R.; Scheller, R.H.; Chavkin, C.; Bajjalieh, S.M. Abnormal neurotransmission in mice lacking synaptic vesicle protein 2A (SV2A). Proc. Natl. Acad. Sci. USA 1999, 96, 15268–15273.
- Custer, K.L.; Austin, N.S.; Sullivan, J.M.; Bajjalieh, S.M. Synaptic vesicle protein 2 enhances release probability at quiescent synapses. J. Neurosci. 2006, 26, 1303–1313.
- Vogl, C.; Tanifuji, S.; Danis, B.; Daniels, V.; Foerch, P.; Wolff, C.; Whalley, B.J.; Mochida, S.; Stephens, G.J. Synaptic vesicle glycoprotein 2A modulates vesicular release and calcium channel function at peripheral sympathetic synapses. Eur. J. Neurosci. 2015, 41, 398–409.
- Xu, T.; Bajjalieh, S.M. SV2 modulates the size of the readily releasable pool of secretory vesicles. Nat. Cell Biol. 2001, 3, 691–698.
- Venkatesan, K.; Alix, P.; Marquet, A.; Doupagne, M.; Niespodziany, I.; Rogister, B.; Seutin, V. Altered balance between excitatory and inhibitory inputs onto CA1 pyramidal neurons from SV2A-deficient but not SV2B-deficient mice. J. Neurosci. Res. 2012, 90, 2317–2327.
- Schivell, A.E.; Mochida, S.; Kensel-Hammes, P.; Custer, K.L.; Bajjalieh, S.M. SV2A and SV2C contain a unique synaptotagmin-binding site. Mol. Cell. Neurosci. 2005, 29, 56–64.
- Yao, J.; Nowack, A.; Kensel-Hammes, P.; Gardner, R.G.; Bajjalieh, S.M. Cotrafficking of SV2 and synaptotagmin at the synapse. J. Neurosci. 2010, 30, 5569–5578.
- Stout, K.A.; Dunn, A.R.; Hoffman, C.; Miller, G.W. The Synaptic Vesicle Glycoprotein 2: Structure, Function, and Disease Relevance. ACS Chem. Neurosci. 2019, 10, 3927–3938.
- Ciruelas, K.; Marcotulli, D.; Bajjalieh, S.M. Synaptic vesicle protein 2: A multi-faceted regulator of secretion. Semin. Cell Dev. Biol. 2019, 95, 130–141.
- Nowack, A.; Malarkey, E.B.; Yao, J.; Bleckert, A.; Hill, J.; Bajjalieh, S.M. Levetiracetam reverses synaptic deficits produced by overexpression of SV2A. PLoS ONE 2011, 6, e29560.
- Meehan, A.L.; Yang, X.; McAdams, B.D.; Yuan, L.L.; Rothman, S.M. A new mechanism for antiepileptic drug action: Vesicular entry may mediate the effects of levetiracetam. J. Neurophysiol. 2011, 106, 1227–1239.
- Mendoza-Torreblanca, J.G.; Vanoye-Carlo, A.; Phillips-Farfán, B.V.; Carmona-Aparicio, L.; Gómez-Lira, G. Synaptic vesicle protein 2A: Basic facts and role in synaptic function. Eur. J. Neurosci. 2013, 38, 3529–3539.
- Nicolas, J.-M.; Hannestad, J.; Holden, D.; Kervyn, S.; Nabulsi, N.; Tytgat, D.; Huang, Y.; Chanteux, H.; Staelens, L.; Matagne, A.; et al. Brivaracetam, a selective high-affinity synaptic vesicle protein 2A (SV2A) ligand with preclinical evidence of high brain permeability and fast onset of action. Epilepsia 2016, 57, 201–209.
- Doheny, H.C.; Ratnaraj, N.; Whittington, M.A.; Jefferys, J.G.; Patsalos, P.N. Blood and cerebrospinal fluid pharmacokinetics of the novel anticonvulsant levetiracetam (ucb L059) in the rat. Epilepsy Res. 1999, 34, 161–168.
- Patsalos, P.N. Pharmacokinetic profile of levetiracetam: Toward ideal characteristics. Pharmacol. Ther. 2000, 85, 77–85.
- Tong, X.; Patsalos, P.N. A microdialysis study of the novel antiepileptic drug levetiracetam: Extracellular pharmacokinetics and effect on taurine in rat brain. Br. J. Pharmacol. 2001, 133, 867–874.
- Kaminski, R.M.; Gillard, M.; Leclercq, K.; Hanon, E.; Lorent, G.; Dassesse, D.; Matagne, A.; Klitgaard, H. Proepileptic phenotype of SV2A-deficient mice is associated with reduced anticonvulsant efficacy of levetiracetam. Epilepsia 2009, 50, 1729–1740.
- De Groot, M.; Aronica, E.; Heimans, J.J.; Reijneveld, J.C. Synaptic vesicle protein 2A predicts response to levetiracetam in patients with glioma. Neurology 2011, 77, 532–539.
- Ohno, Y.; Ishihara, S.; Terada, R.; Kikuta, M.; Sofue, N.; Kawai, Y.; Serikawa, T.; Sasa, M. Preferential increase in the hippocampal synaptic vesicle protein 2A (SV2A) by pentylenetetrazole kindling. Biochem. Biophys. Res. Commun. 2009.
- Matveeva, E.A.; Vanaman, T.C.; Whiteheart, S.W.; Slevin, J.T. Levetiracetam prevents kindling-induced asymmetric accumulation of hippocampal 7S SNARE complexes. Epilepsia 2008, 49, 1749–1758.
- Inaba, T.; Miyamoto, N.; Hira, K.; Ueno, Y.; Yamashiro, K.; Watanabe, M.; Shimada, Y.; Hattori, N.; Urabe, T. Protective Role of Levetiracetam Against Cognitive Impairment And Brain White Matter Damage in Mouse prolonged Cerebral Hypoperfusion. Neuroscience 2019, 414, 255–264.
- Contreras-García, I.J.; Gómez-Lira, G.; Phillips-Farfán, B.V.; Pichardo-Macías, L.A.; García-Cruz, M.E.; Chávez-Pacheco, J.L.; Mendoza-Torreblanca, J.G. Synaptic Vesicle Protein 2A Expression in Glutamatergic Terminals Is Associated with the Response to Levetiracetam Treatment. Brain Sci. 2021, 11, 531.
- Marcotulli, D.; Fattorini, G.; Bragina, L.; Perugini, J.; Conti, F. Levetiracetam Affects Differentially Presynaptic Proteins in Rat Cerebral Cortex. Front. Cell. Neurosci. 2017, 11, 389.
- Niespodziany, I.; Klitgaard, H.; Margineanu, D.G. Levetiracetam inhibits the high-voltage-activated Ca(2+) current in pyramidal neurones of rat hippocampal slices. Neurosci. Lett. 2001, 306, 5–8.
- Costa, C.; Martella, G.; Picconi, B.; Prosperetti, C.; Pisani, A.; Di Filippo, M.; Pisani, F.; Bernardi, G.; Calabresi, P. Multiple mechanisms underlying the neuroprotective effects of antiepileptic drugs against in vitro ischemia. Stroke 2006, 37, 1319–1326.
- Pisani, A.; Bonsi, P.; Martella, G.; De Persis, C.; Costa, C.; Pisani, F.; Bernardi, G.; Calabresi, P. Intracellular calcium increase in epileptiform activity: Modulation by levetiracetam and lamotrigine. Epilepsia 2004, 45, 719–728.
- Lukyanetz, E.A.; Shkryl, V.M.; Kostyuk, P.G. Selective blockade of N-type calcium channels by levetiracetam. Epilepsia 2002, 43, 9–18.
- Yan, H.-D.; Ishihara, K.; Seki, T.; Hanaya, R.; Kurisu, K.; Arita, K.; Serikawa, T.; Sasa, M. Inhibitory effects of levetiracetam on the high-voltage-activated L-type Ca2+ channels in hippocampal CA3 neurons of spontaneously epileptic rat (SER). Brain Res. Bull. 2013, 90, 142–148.
- Deshpande, L.S.; Delorenzo, R.J. Mechanisms of levetiracetam in the control of status epilepticus and epilepsy. Front. Neurol. 2014, 5, 11.
- Vogl, C.; Mochida, S.; Wolff, C.; Whalley, B.J.; Stephens, G.J. The synaptic vesicle glycoprotein 2A ligand levetiracetam inhibits presynaptic Ca2+ channels through an intracellular pathway. Mol. Pharmacol. 2012, 82, 199–208.
- Angehagen, M.; Margineanu, D.G.; Ben-Menachem, E.; Rönnbäck, L.; Hansson, E.; Klitgaard, H. Levetiracetam reduces caffeine-induced Ca2+ transients and epileptiform potentials in hippocampal neurons. Neuroreport 2003, 14, 471–475.
- Nagarkatti, N.; Deshpande, L.S.; DeLorenzo, R.J. Levetiracetam inhibits both ryanodine and IP3 receptor activated calcium induced calcium release in hippocampal neurons in culture. Neurosci. Lett. 2008, 436, 289–293.
- Cataldi, M.; Lariccia, V.; Secondo, A.; di Renzo, G.; Annunziato, L. The antiepileptic drug levetiracetam decreases the inositol 1,4,5-trisphosphate-dependent I increase induced by ATP and bradykinin in PC12 cells. J. Pharmacol. Exp. Ther. 2005, 313, 720–730.
- Navidhamidi, M.; Ghasemi, M.; Mehranfard, N. Epilepsy-associated alterations in hippocampal excitability. Rev. Neurosci. 2017, 28, 307–334.
- Rigo, J.-M.; Hans, G.; Nguyen, L.; Rocher, V.; Belachew, S.; Malgrange, B.; Leprince, P.; Moonen, G.; Selak, I.; Matagne, A.; et al. The anti-epileptic drug levetiracetam reverses the inhibition by negative allosteric modulators of neuronal GABA- and glycine-gated currents. Br. J. Pharmacol. 2002, 136, 659–672.
- Doelken, M.T.; Hammen, T.; Bogner, W.; Mennecke, A.; Stadlbauer, A.; Boettcher, U.; Doerfler, A.; Stefan, H. Alterations of intracerebral γ-aminobutyric acid (GABA) levels by titration with levetiracetam in patients with focal epilepsies. Epilepsia 2010, 51, 1477–1482.
- Li, Q.; Chen, C.; Gong, T. High-field MRS study of GABA+ in patients with migraine: Response to levetiracetam treatment. Neuroreport 2018, 29, 1007–1010.
- Klitgaard, H.; Matagne, A.; Grimee, R.; Vanneste-Goemaere, J.; Margineanu, D.G. Electrophysiological, neurochemical and regional effects of levetiracetam in the rat pilocarpine model of temporal lobe epilepsy. Seizure 2003, 12, 92–100.
- Fukuyama, K.; Tanahashi, S.; Nakagawa, M.; Yamamura, S.; Motomura, E.; Shiroyama, T.; Tanii, H.; Okada, M. Levetiracetam inhibits neurotransmitter release associated with CICR. Neurosci. Lett. 2012, 518, 69–74.
- Pichardo Macías, L.A.; Ramírez Mendiola, B.A.; Contreras García, I.J.; Zamudio Hernández, S.R.; Chávez Pacheco, J.L.; Sánchez Huerta, K.B.; Mendoza Torreblanca, J.G. Effect of levetiracetam on extracellular amino acid levels in the dorsal hippocampus of rats with temporal lobe epilepsy. Epilepsy Res. 2018, 140, 111–119.
- Löscher, W.; Hönack, D.; Bloms-Funke, P. The novel antiepileptic drug levetiracetam (ucb L059) induces alterations in GABA metabolism and turnover in discrete areas of rat brain and reduces neuronal activity in substantia nigra pars reticulata. Brain Res. 1996, 735, 208–216.
- Mazzuferi, M.; Palma, E.; Martinello, K.; Maiolino, F.; Roseti, C.; Fucile, S.; Fabene, P.F.; Schio, F.; Pellitteri, M.; Sperk, G.; et al. Enhancement of GABA(A)-current run-down in the hippocampus occurs at the first spontaneous seizure in a model of temporal lobe epilepsy. Proc. Natl. Acad. Sci. USA 2010, 107, 3180–3185.
- Cifelli, P.; Palma, E.; Roseti, C.; Verlengia, G.; Simonato, M. Changes in the sensitivity of GABAA current rundown to drug treatments in a model of temporal lobe epilepsy. Front. Cell. Neurosci. 2013, 7, 108.
- Palma, E.; Ragozzino, D.; Di Angelantonio, S.; Mascia, A.; Maiolino, F.; Manfredi, M.; Cantore, G.; Esposito, V.; Di Gennaro, G.; Quarato, P.; et al. The antiepileptic drug levetiracetam stabilizes the human epileptic GABAA receptors upon repetitive activation. Epilepsia 2007, 48, 1842–1849.
- Malatynska, E.; Knapp, R.; Ikeda, M.; Yamamura, H.I. Beta-carboline interactions at the BZ-GABA receptor chloride-ionophore complex in the rat cerebral cortex. Brain Res. Bull. 1989, 22, 845–848.
- Evans, A.K.; Lowry, C.A. Pharmacology of the beta-carboline FG-7,142, a partial inverse agonist at the benzodiazepine allosteric site of the GABA A receptor: Neurochemical, neurophysiological, and behavioral effects. CNS Drug Rev. 2007, 13, 475–501.
- Kulick, C.V.; Gutherz, S.B.; Beck, V.C.; Medvedeva, N.; Soper, C.; Forcelli, P.A. Profile of anticonvulsant action of levetiracetam, tiagabine and phenobarbital against seizures evoked by DMCM (methyl-6,7-dimethoxy-4-ethyl-β-carboline-3-carboxylate) in neonatal rats. Eur. J. Pharmacol. 2014, 743, 63–68.
- Wakita, M.; Kotani, N.; Kogure, K.; Akaike, N. Inhibition of excitatory synaptic transmission in hippocampal neurons by levetiracetam involves Zn2+-dependent GABA type A receptor-mediated presynaptic modulation. J. Pharmacol. Exp. Ther. 2014, 348, 246–259.
- Buckley, K.; Kelly, R.B. Identification of a transmembrane glycoprotein specific for secretory vesicles of neural and endocrine cells. J. Cell Biol. 1985, 100, 1284–1294.
- Bajjalieh, S.M.; Frantz, G.D.; Weimann, J.M.; McConnell, S.K.; Scheller, R.H. Differential expression of synaptic vesicle protein 2 (SV2) isoforms. J. Neurosci. 1994, 14, 5223–5235.
- Bragina, L.; Fattorini, G.; Giovedí, S.; Melone, M.; Bosco, F.; Benfenati, F.; Conti, F. Analysis of Synaptotagmin, SV2, and Rab3 Expression in Cortical Glutamatergic and GABAergic Axon Terminals. Front. Cell. Neurosci. 2011, 5, 32.
- Grønborg, M.; Pavlos, N.J.; Brunk, I.; Chua, J.J.E.; Münster-Wandowski, A.; Riedel, D.; Ahnert-Hilger, G.; Urlaub, H.; Jahn, R. Quantitative comparison of glutamatergic and GABAergic synaptic vesicles unveils selectivity for few proteins including MAL2, a novel synaptic vesicle protein. J. Neurosci. 2010, 351, 981–984.
- Tokudome, K.; Okumura, T.; Terada, R.; Shimizu, S.; Kunisawa, N.; Mashimo, T.; Serikawa, T.; Sasa, M.; Ohno, Y. A Missense Mutation of the Gene Encoding Synaptic Vesicle Glycoprotein 2A (SV2A) Confers Seizure Susceptibility by Disrupting Amygdalar Synaptic GABA Release. Front. Pharmacol. 2016, 7, 210.
- Ohno, Y.; Tokudome, K. Therapeutic Role of Synaptic Vesicle Glycoprotein 2A (SV2A) in Modulating Epileptogenesis. CNS Neurol. Disord. Drug Targets 2017, 16, 463–471.
- Contreras-García, I.J.; Pichardo-Macías, L.A.; Santana-Gómez, C.E.; Sánchez-Huerta, K.; Ramírez-Hernández, R.; Gómez-González, B.; Rocha, L.; Mendoza Torreblanca, J.G. Differential expression of synaptic vesicle protein 2A after status epilepticus and during epilepsy in a lithium-pilocarpine model. Epilepsy Behav. 2018, 88, 283–294.
- Tokudome, K.; Okumura, T.; Shimizu, S.; Mashimo, T.; Takizawa, A.; Serikawa, T.; Terada, R.; Ishihara, S.; Kunisawa, N.; Sasa, M.; et al. Synaptic vesicle glycoprotein 2A (SV2A) regulates kindling epileptogenesis via GABAergic neurotransmission. Sci. Rep. 2016, 6, 27420.
- Mendoza-Torreblanca, J.G.; García-Cruz, M.E.; Sánchez-Cruz, I.; Gomez-Gonzalez, B.; Juárez-Méndez, S.; Gómez-Lira, G. Analysis of Differential Expression of Synaptic Vesicle Protein 2A in the Adult Rat Brain. Neuroscience 2019, 419, 108–120.
- Micov, A.; Tomić, M.; Popović, B.; Stepanović-Petrović, R. The antihyperalgesic effect of levetiracetam in an inflammatory model of pain in rats: Mechanism of action. Br. J. Pharmacol. 2010, 161, 384–392.
- Stepanović-Petrović, R.M.; Micov, A.M.; Tomić, M.A.; Ugrešić, N.D. The local peripheral antihyperalgesic effect of levetiracetam and its mechanism of action in an inflammatory pain model. Anesth. Analg. 2012, 115, 1457–1466.
- Carunchio, I.; Pieri, M.; Ciotti, M.T.; Albo, F.; Zona, C. Modulation of AMPA receptors in cultured cortical neurons induced by the antiepileptic drug levetiracetam. Epilepsia 2007, 48, 654–662.
- Hentschke, M.; Wiemann, M.; Hentschke, S.; Kurth, I.; Hermans-Borgmeyer, I.; Seidenbecher, T.; Jentsch, T.J.; Gal, A.; Hübner, C.A. Mice with a targeted disruption of the Cl-/HCO3- exchanger AE3 display a reduced seizure threshold. Mol. Cell. Biol. 2006, 26, 182–191.
- Svichar, N.; Esquenazi, S.; Chen, H.-Y.; Chesler, M. Preemptive regulation of intracellular pH in hippocampal neurons by a dual mechanism of depolarization-induced alkalinization. J. Neurosci. 2011, 31, 6997–7004.
- Sander, T.; Toliat, M.R.; Heils, A.; Leschik, G.; Becker, C.; Rüschendorf, F.; Rohde, K.; Mundlos, S.; Nürnberg, P. Association of the 867Asp variant of the human anion exchanger 3 gene with common subtypes of idiopathic generalized epilepsy. Epilepsy Res. 2002, 51, 249–255.
- Bonnet, U.; Bingmann, D.; Speckmann, E.-J.; Wiemann, M. Levetiracetam mediates subtle pH-shifts in adult human neocortical pyramidal cells via an inhibition of the bicarbonate-driven neuronal pH-regulation—Implications for excitability and plasticity modulation. Brain Res. 2019, 1710, 146–156.
- Leniger, T.; Thöne, J.; Bonnet, U.; Hufnagel, A.; Bingmann, D.; Wiemann, M. Levetiracetam inhibits Na+-dependent Cl-/HCO3- exchange of adult hippocampal CA3 neurons from guinea-pigs. Br. J. Pharmacol. 2004, 142, 1073–1080.
- Gu, J.; Lynch, B.A.; Anderson, D.; Klitgaard, H.; Lu, S.; Elashoff, M.; Ebert, U.; Potschka, H.; Löscher, W. The antiepileptic drug levetiracetam selectively modifies kindling-induced alterations in gene expression in the temporal lobe of rats. Eur. J. Neurosci. 2004, 19, 334–345.
- Husum, H.; Bolwig, T.G.; Sánchez, C.; Mathé, A.A.; Hansen, S.L. Levetiracetam prevents changes in levels of brain-derived neurotrophic factor and neuropeptide Y mRNA and of Y1- and Y5-like receptors in the hippocampus of rats undergoing amygdala kindling: Implications for antiepileptogenic and mood-stabilizing proper. Epilepsy Behav. 2004, 5, 204–215.
- Christensen, K.V.; Leffers, H.; Watson, W.P.; Sánchez, C.; Kallunki, P.; Egebjerg, J. Levetiracetam attenuates hippocampal expression of synaptic plasticity-related immediate early and late response genes in amygdala-kindled rats. BMC Neurosci. 2010, 11, 9.
- Kim, J.-E.; Choi, H.-C.; Song, H.-K.; Jo, S.-M.; Kim, D.-S.; Choi, S.-Y.; Kim, Y.-I.; Kang, T.-C. Levetiracetam inhibits interleukin-1 beta inflammatory responses in the hippocampus and piriform cortex of epileptic rats. Neurosci. Lett. 2010, 471, 94–99.
- Rassu, M.; Biosa, A.; Galioto, M.; Fais, M.; Sini, P.; Greggio, E.; Piccoli, G.; Crosio, C.; Iaccarino, C. Levetiracetam treatment ameliorates LRRK2 pathological mutant phenotype. J. Cell. Mol. Med. 2019, 23, 8505–8510.
- Kovacevic, J.; Maroteaux, G.; Schut, D.; Loos, M.; Dubey, M.; Pitsch, J.; Remmelink, E.; Koopmans, B.; Crowley, J.; Cornelisse, L.N.; et al. Protein instability, haploinsufficiency, and cortical hyper-excitability underlie STXBP1 encephalopathy. Brain 2018, 141, 1350–1374.
- Dilena, R.; Striano, P.; Traverso, M.; Viri, M.; Cristofori, G.; Tadini, L.; Barbieri, S.; Romeo, A.; Zara, F. Dramatic effect of levetiracetam in early-onset epileptic encephalopathy due to STXBP1 mutation. Brain Dev. 2016, 38, 128–131.
- Parveen, B.; Tripathi, M.; Vohora, D. A Cross-Sectional Study to Assess the Modulation of Wnt Inhibitors following Anti-Epileptic Drug Therapy and their Correlation with Vitamin D and Receptor Activator of Nuclear Factor κ B Ligand in Indian Women with Epilepsy. Basic Clin. Pharmacol. Toxicol. 2018, 123, 271–276.
- Lange, F.; Weßlau, K.; Porath, K.; Hörnschemeyer, J.; Bergner, C.; Krause, B.J.; Mullins, C.S.; Linnebacher, M.; Köhling, R.; Kirschstein, T. AMPA receptor antagonist perampanel affects glioblastoma cell growth and glutamate release in vitro. PLoS ONE 2019, 14, e0211644.
- Niidome, K.; Taniguchi, R.; Yamazaki, T.; Tsuji, M.; Itoh, K.; Ishihara, Y. FosL1 Is a Novel Target of Levetiracetam for Suppressing the Microglial Inflammatory Reaction. Int. J. Mol. Sci. 2021, 22, 10962.
- Hassel, B.; Taubøll, E.; Shaw, R.; Gjerstad, L.; Dingledine, R. Region-specific changes in gene expression in rat brain after chronic treatment with levetiracetam or phenytoin. Epilepsia 2010, 51, 1714–1720.
- Sills, G.J.; Rogawski, M.A. Mechanisms of action of currently used antiseizure drugs. Neuropharmacology 2020, 168, 107966.
- Belcastro, V.; Pierguidi, L.; Tambasco, N. Levetiracetam in brain ischemia: Clinical implications in neuroprotection and prevention of post-stroke epilepsy. Brain Dev. 2011, 33, 289–293.
- Rakhade, S.N.; Shah, A.K.; Agarwal, R.; Yao, B.; Asano, E.; Loeb, J.A. Activity-dependent gene expression correlates with interictal spiking in human neocortical epilepsy. Epilepsia 2007, 48 (Suppl. 5), 86–95.
- Arion, D.; Sabatini, M.; Unger, T.; Pastor, J.; Alonso-Nanclares, L.; Ballesteros-Yáñez, I.; García Sola, R.; Muñoz, A.; Mirnics, K.; DeFelipe, J. Correlation of transcriptome profile with electrical activity in temporal lobe epilepsy. Neurobiol. Dis. 2006, 22, 374–387.
- Margineanu, D.G.; Matagne, A.; Kaminski, R.M.; Klitgaard, H. Effects of chronic treatment with levetiracetam on hippocampal field responses after pilocarpine-induced status epilepticus in rats. Brain Res. Bull. 2008, 77, 282–285.
- Zhao, T.; Yu, J.; Wang, T.-T.; Feng, J.; Zhao, W.-B.; Sun, L.; Yu, L.-H.; Li, H.-J.; Sun, Y. Impact of ABCB1 Polymorphism on Levetiracetam Serum Concentrations in Epileptic Uygur Children in China. Ther. Drug Monit. 2020, 42, 886–892.
- Calame, D.G.; Herman, I.; Riviello, J.J. A de novo heterozygous rare variant in SV2A causes epilepsy and levetiracetam-induced drug-resistant status epilepticus. Epilepsy Behav. Rep. 2021, 15, 100425.
- Wolking, S.; Moreau, C.; Nies, A.T.; Schaeffeler, E.; McCormack, M.; Auce, P.; Avbersek, A.; Becker, F.; Krenn, M.; Møller, R.S.; et al. Testing association of rare genetic variants with resistance to three common antiseizure medications. Epilepsia 2020, 61, 657–666.
- Grimminger, T.; Pernhorst, K.; Surges, R.; Niehusmann, P.; Priebe, L.; von Lehe, M.; Hoffmann, P.; Cichon, S.; Schoch, S.; Becker, A.J. Levetiracetam resistance: Synaptic signatures & corresponding promoter SNPs in epileptic hippocampi. Neurobiol. Dis. 2013, 60, 115–125.
- Helmstaedter, C.; Mihov, Y.; Toliat, M.R.; Thiele, H.; Nuernberg, P.; Schoch, S.; Surges, R.; Elger, C.E.; Kunz, W.S.; Hurlemann, R. Genetic variation in dopaminergic activity is associated with the risk for psychiatric side effects of levetiracetam. Epilepsia 2013, 54, 36–44.