Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Camila Xu and Version 1 by Ken Mix.
Silicon (Si) is present in soil mainly in three different phases such as solid, liquid, and adsorbed. Solid phases can be either amorphous or crystalline. Plants take up silicon (Si)Si from the soil which impacts their growth and nutrient accumulation. It increases plant resistance to abiotic and biotic stresses such as drought, salinity, and heavy metal, diseases, and pest infestation.
- growth
- nutrients
- drought
- silicon
- plants
1. Introduction
Most of the major crops accumulate a significant amount of silicon (Si) though it is considered non-essential for plant growth and development [1]. While Si does not directly contribute to plant metabolism [2[2][3],3], there is evidence that Si improves: crop productivity [4[4][5][6][7],5,6,7], nutrient accumulation in plants [8,9,10[8][9][10][11],11], heavy metal resistance in grain crops [12[12][13][14],13,14], and increases drought tolerance of plants [15,16,17][15][16][17]. Since these benefits for crop production have been recognized, the global use of Si as a soil amendment is increasing.
The Earth’s pedosphere of Si is estimated to be 28.2% by weight [3]. Si, along with oxygen and metals, forms silicon dioxide (Si2O) and water-soluble silicates. Si minerals go through various physical and chemical weathering and release Si in solution under suitable pH condition. The source of silica and silicates in soil and clays is from the weathered Si minerals such as quartz and feldspar present in the pedosphere [18,19][18][19]. Si is present in soil mainly in three different phases such as solid, liquid, and adsorbed. Solid phases can be either amorphous or crystalline. Amorphous silica contributes significantly more to dissolve Si in soil solution because of its higher solubility than the crystalline form [3]. However, the plant does not uptake any Si as amorphous silica; rather it is taken up by plants in the form of monosilicic acid (H4SiO4) [20].
The addition of Si as a soil amendment in the crop field is a recent phenomenon, and few reports have been published on the impact of Si on plants. The recent publications report that Si affects growth, yield [21,22[21][22][23],23], and disease resistance [24,25,26,27][24][25][26][27] among other plant conditions.
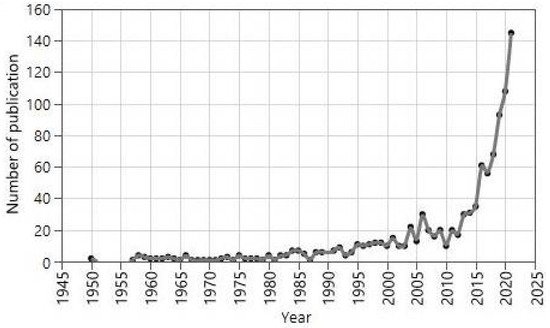
Figure 1 shows the number of publications vs. year for the time period 1950–2021. The research on the plant silicon relationship began after 1935. Until 1980, research on this field was minimal. However, between 1980–2000, the overall number of publications started to increase gradually. After 2010, a dramatic increase in publication indicates that this field is getting more research attention [28].

It is noteworthy, that among the ten major produced crops in the world, seven crops (rice, wheat, sugarcane, sugar beet, soybean, tomato, and barley) are Si accumulators and their biomasses consist of more than 1% Si in dry matter [29]. Though the ability of Si accumulation in these plants highly varies [30], production of these crops takes away large amounts of Si from the soil. For instance, rice and sugarcane individually can remove approximately 500 kg and 300 kg of Si per ha, respectively, while harvesting [20,31][20][31]. After several years of continuous cropping and harvesting of plants, available Si declines in the soil. This fact necessitates the application of Si from external sources. In different parts of the world, such as India, Vietnam, China, researchers used rice [32], wheat [17], and sugarcane [20] for experiments on Si applications related to crop production. It has been observed that Si has a beneficial effect, including increasing crop yield, limiting abiotic stresses from salinity, increasing drought tolerance [33], and reducing the toxic effects [34,35][34][35] of heavy metals. [36]. The presence of higher Si in the plants’ roots, shoots, and leaves makes the plants more resistant to pest attack and enhances drought tolerance as it forms a thick layer under the cuticle. This Si layer reduces leaf digestibility for pests as well as water transpiration loss from plants [37]. Silicate application sharply decreased transpirational flow in rice about 4.2% to 0.8% [38]. It was reported that applications of amorphous silica minimize the cadmium stress in the plants by inhibiting root to shoot transfer of cadmium along with other metals [39,40][39][40]. When amorphous silica was applied at a rate of 1000 kg/ha, it increased the availability and accumulation of mineral nutrients: P (10–40%), Ca (up to 33%), S (up to 51%), Mo (up to 54%), and Cu (10–40%) [41]. Si helps make the nutrients available to plant root systems by impeding soil particles from bonding with mineral nutrients. It is known that the content of Si in agricultural soils is declining, both due to natural weathering and continuous agricultural and activities [42].
2. Role of Si on Plant Growth and Yield
Application of Si-based fertilizer has been reported to be effective for plant growth and yield. To determine the effect of Si fertilizer on growth, yield, and nutrient accumulation of the rice plant, four different dosages of SiO2 were applied with identical recommended dosages of N, P, and K fertilizers. It was observed that yield components (number of tillers, number of panicles per plant, and number of grains per panicle) and yield were significantly affected by Si fertilizer doses. About 3716 kg/ha maximum grain yield was obtained when they applied SiO2 at a rate of 329 kg/ha. There was about a 23% increase in grain yield compared to the control [8]. Whereas in a similar study with different Si fertilizer (Na2SiO3) dosages, up to 17.4% yield increase along with increased panicle numbers were recorded from a field trial in China. The increased growth and yield recorded from this experiment are given in Table 1.Table 1. Effect of Si fertilization on rice growth and yield in China [4].
In India, researchers used diatomaceous earth (DE) as a source of Si and compared its use in two different moisture regimes: saturated/submerged and field capacity. It was found that biomass yield was high with almost all of the DE treatments in acidic (300 and 600 kg/ha) and alkaline (150, 300 and 600 kg/ha) soil condition. Analysis of soil and rice yield, before and after the application of DE, provided evidence that applications of DE increase rice yield regardless of the soil condition. The increases were 150, 300, and 600 kg/ha in alkaline, acidic, and neutral soil, respectively. According to the report, DE works best in submerged condition compared to field capacity condition of the rice field [4]. Si is also responsible for grain quality in rice. Formation of the quality hull with milky sap is high when the concentration of Si in rice shoot is high [43].
Si fertilization has a positive impact on wheat production. It increases the plant height, no. of spikelets, and number of spikes per spikelet. In irrigated fields, grain yield increased by 13.4% compared to no Si application. It has been reported that the application of K2SiO3, at a rate of 12 kg/ha, increased plant height, the number of effective tillers per m2 up to 515.33, spike length up to 12.25 cm, and number of spikelets per spike on an average 16.70. A maximum grain yield of 4380 kg/ha was observed when K2SiO3 was applied with four irrigations [17]. Exogenous application of Si or a combination of Si and Se were reported to change the physio-biochemical activity in wheat which resulted in the successful growth of wheat in saline soil [44]. Foliar application of sodium silicate salt (Na2Si3O7), especially at the tillering stage and anthesis stage, increases the yield for various wheat cultivars [45]. In contrast to other materials, volcanic tuff (Chem comp) was applied in Idaho and showed no remarkable effect. There is also evidence that these improvements in yield are only related to stressed conditions and that no improvement in yield is observed during ideal growing conditions [46].
Notable improvements were observed when Si fertilizer was applied for maize production [47]. In Pakistan, two hybrid maize varieties, P-33H25 and FH-810, were grown under 100% and 60% field capacity of water levels. In a water deficit condition, application of Si significantly increased plant height, stem diameter, and cob length (13.96 cm and 12.83 cm, respectively) for both maize varieties. It also increases the number of grains per cob (235.05 and 215.35) and grain yield (0.46 kg and 0.39 kg) correspondingly. This increased yield was due to the increased number of cobs, grains per cob, and weight of 1000 grains [7]. A similar result in the drought stressed condition was published on the improvement of maize plant growth and yield from Turkey and Greenville, South Carolina [22,48][22][48].
Si is not only beneficial for cereal and grain crops; it also increases the production and fresh weight of vegetables. In India, studies were conducted from 2013 to 2017 on three different potato varieties. Application of additional Si (ferti-silica 50 mg/dm3) increased tuber yield by 15–50% [21]. In a pot cultivation system, Ca and Mg silicate were used for growing potato in the absence and presence of water. According to this report, Si application enhances Si availability in soil, which increases overall tuber dry weight irrespective of water condition. Though there was no significant improvement on an increase in the number of tubers/plants [49].
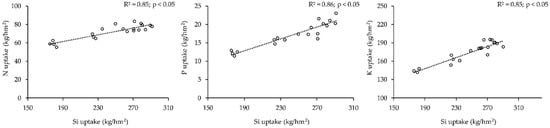
 Figure 2. Linear regression between Si uptake and nitrogen (N), phosphorus (P), and potassium (K) uptakes in above-ground biomass of rice variety BC15 [8].
There appears to be evidence that very high doses of Si decreased the net accumulation of N and P in the plant’s shoots and roots, though it is thought that the concentration of N decreased due to the increased growth rate and decreased Mg availability caused by Si treatment [42,62][42][56]. Silicon deposition in endodermal cells of plant roots, which may contribute to decreased P uptake, is one probable reason for the decrease in P accumulation. The generation of apoplastic barriers to P permeability across roots produced by Si deposition in roots, which reduces P uptake, is primarily responsible for this effect [63,64][57][58]. Another concept is that the creation of a cuticle-silica double layer in leaves as a result of Si deposition lowers the plant’s transpiration rate. Transpiration is negatively connected with the Si content of rice aerial parts, and when the SiO2 concentration of shoots surpasses 10% of the dry matter weight, the rice transpiration rate might be lowered by 20–30% [65][59]. In contrast, another study found Si application does not have any measurable effect on increasing extractable phosphorus from soil, in fact it may increase P fixation in soil because Si increases soil pH which influences soil P to be strongly adsorbed by soil particles and silicic acid is not strong enough to break that bond [66][60]. Therefore, the supplemental Si application slightly affects the availability of P.
Figure 2. Linear regression between Si uptake and nitrogen (N), phosphorus (P), and potassium (K) uptakes in above-ground biomass of rice variety BC15 [8].
There appears to be evidence that very high doses of Si decreased the net accumulation of N and P in the plant’s shoots and roots, though it is thought that the concentration of N decreased due to the increased growth rate and decreased Mg availability caused by Si treatment [42,62][42][56]. Silicon deposition in endodermal cells of plant roots, which may contribute to decreased P uptake, is one probable reason for the decrease in P accumulation. The generation of apoplastic barriers to P permeability across roots produced by Si deposition in roots, which reduces P uptake, is primarily responsible for this effect [63,64][57][58]. Another concept is that the creation of a cuticle-silica double layer in leaves as a result of Si deposition lowers the plant’s transpiration rate. Transpiration is negatively connected with the Si content of rice aerial parts, and when the SiO2 concentration of shoots surpasses 10% of the dry matter weight, the rice transpiration rate might be lowered by 20–30% [65][59]. In contrast, another study found Si application does not have any measurable effect on increasing extractable phosphorus from soil, in fact it may increase P fixation in soil because Si increases soil pH which influences soil P to be strongly adsorbed by soil particles and silicic acid is not strong enough to break that bond [66][60]. Therefore, the supplemental Si application slightly affects the availability of P.
| Application Rate (kg/ha) |
Number of Panicles (×104/ha) |
Number of Spikelets/Panicle | Yield (kg/ha) |
|---|---|---|---|
| 0 | 4.84 | 74.7 | 7010 |
| 75 | 4.94 | 73.9 | 7870 |
| 105 | 5.03 | 74.8 | 8160 |
| 135 | 5.03 | 76.8 | 8230 |
3. Role of Si on Nutrients Availability and Accumulation
There are sixteen minerals playing essential roles in the plant’s cell metabolism, energy transfer, osmosis, and reproduction. Among these sixteen elements, nitrogen (N) is a major constituent of the plant structure which works with a combination of H, C, and P. Si mostly affects the availability of N in soil and N accumulation in plants [55][50]. It forms the organic compounds such as protein and nucleotide in the plant. Similar to N, phosphorus (P) also forms some organophosphorus compound such as sugar phosphate, pyrophosphate bond (ATP), phytin, etc. On the other hand, potassium (K) maintains the ionic balance among the cells along with the activation of enzymes. It also provides mechanical strength against the lodging of plants in water deficit condition. Among the micronutrients, Fe and Mn work as a cofactor of enzymes and help in N metabolism.3.1. Macronutrients
In the published literature, the results of Si application reveal a mixed trend regarding macronutrient accumulation. Needless to say, the environmental parameters, Si dosage, and soil type of the research sites were widely variable. Therefore, one cannot argue the results are contradictory. Some reports claim the improvement of nutrient accumulation by using silicon. According to Cuong et al. [8] (Figure 2), the application of silica has a positive impact on almost all the aspects such as availability, assimilation, and uptake of N, P, and K in rice plants, especially in grains [57,58][51][52]. The increase in N, P, and K accumulation recorded up to 33%, 69%, and 36.8%, respectively, compared to the control [8]. Similar results were also found utilizing diatomaceous earth on the rice field [58][52]. It was concluded in another study that Si fertilization has a positive correlation with P uptake making P more available in soil [59][53]. Generally, P concentration increases in root areas but in potato a higher concentration of P was found in the leaves due to the application of Ca and Mg silicate fertilizers [60][54]. The plant’s available forms of phosphorus also increased in soil because Si binds with iron and manganese, thus preventing phosphorus opportunity to bond with those elements [49]. Whereas potassium concentration in the shoots and roots decreased in lettuce due to the addition of Si and increased for some other crops such as maize and rice from 10–40% [42]. Hence, the accumulation of potassium with the addition of Si appears to be dependent on plant species. In addition to increased N accumulation, S and Mg accumulation also increased in total plant biomass with a high concentration in roots when Si treatments continued for a longer duration (3 weeks). Though, a very high dose of Si decreases the availability of Mg [42,61][42][55].
Figure 2. Linear regression between Si uptake and nitrogen (N), phosphorus (P), and potassium (K) uptakes in above-ground biomass of rice variety BC15 [8].
3.2. Micronutrients
Very limited reports are available on Si applications associated with micronutrient accumulation; however, Si application appears to have an impact on crop accumulation of most of the micronutrients. The net accumulation of Fe and Mn has been improved by Si application as well as boron accumulation in plant leaves. Fe concentration increased in both the roots and shoots, respectively, 20–40% and 10% [43]. While Si did not influence the accumulation of Cl and Mo, it decreased the accumulation of Cu and Zn by 20%. [10]. In numerous studies, Si has been shown to limit Zn bioavailability in soil by redistributing the metal to more stable fractions such as organic materials and crystalline Fe oxides [67][61]. Silicon impacts the exudation of several organic acids (e.g., oxalic, acetic, tartaric, maleic, and fumaric acids) from rice roots, which may be involved in Zn toxicity mitigation via immobilization/co-precipitation in the soil solution [68][62]. In general, the immobilization of harmful Cu ions by increased cell wall binding capacity and the creation of Cu-binding molecules, both in roots and shoots, is attributed to Si-mediated reduction in Cu toxicity. Si lowered the expression of two Cu transporter genes in Arabidopsis roots, AtCOPT1 and AtHMA5. According to the authors, Si deposits generated in cell walls boosted Cu-binding sites, reducing the impact of elevated Cu levels in plant cells [69][63]. According to Flora et al. [70][64], the Si-mediated reduction in Cu toxicity in tobacco (Nicotiana tabacum) via decreasing root uptake of Cu also lowers the expression of NtCOPT1 and raises the expression of genes involved in ethylene production. Kim et al. [71][65] proposed that increased Si buildup in the roots of Cu-stressed rice plants inhibited Cu influx through down-regulating metal transporter genes OsHMA2 and OsHMA3. In many cases, the combined application of Si with other minerals such as Zn increases the availability and accumulation of micronutrients. Zn concentration increased up to 10 µg/L in all organs of rice plants when additional Zn was applied in combination with Si fertilizers. On the other hand, in Zn deficient conditions, Si application increases the Ca concentration in rice and maize shoots and grains [10,22][10][22]. In boron (B)-deficient conditions, Si increases Zn, Mo, Mn, and Cu in sunflower shoots. However, it decreases Fe concentration in roots but increases Fe in fully developed leaves, increasing its mobility [72][66].3.3. Silicon
Increased Si availability in soil solution is the main reason for increased Si uptake in plants. Additional Si application stimulates Si uptake by plants as it improves the root system. Si uptake and accumulation are highest for Si accumulator plants, which consist of more than 1% of total biomass silicon [73][67]. In Table 2, a few major crops are listed with Si percentage in total biomass. Rice plants are the highest Si accumulators, followed by wheat, barley, tomato, and sugarcane. In rice cultivation systems, the application of SiO2 at a rate of 100–400 kg/ha increases Si uptake 26.8–58.5% in total plant biomass [8].Table 2. Si% in above-ground parts of major crop plants [74].
In potato, the soil and foliar application of sodium metasilicate have different effects on the leaf, stem, and tuber Si concentration, and accumulation. The concentration of Si was maximum in stems for soil-applied Si compared to foliar-applied Si and untreated control. No significant differences were found in tuber Si concentration. Whereas Si accumulation in the stem was maximum for foliar application of Si. Overall, soil application of Si provides maximum Si concentration and accumulation in different parts of the potato plant [38]. Figure 3 shows the comparison of Si concentration and accumulation in leaves, roots, stems, and tubers of potato for soil- and foliar-applied sodium metasilicate.
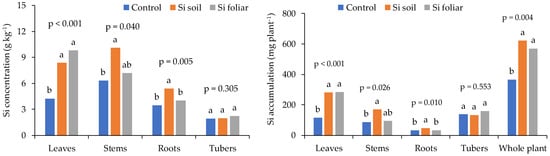
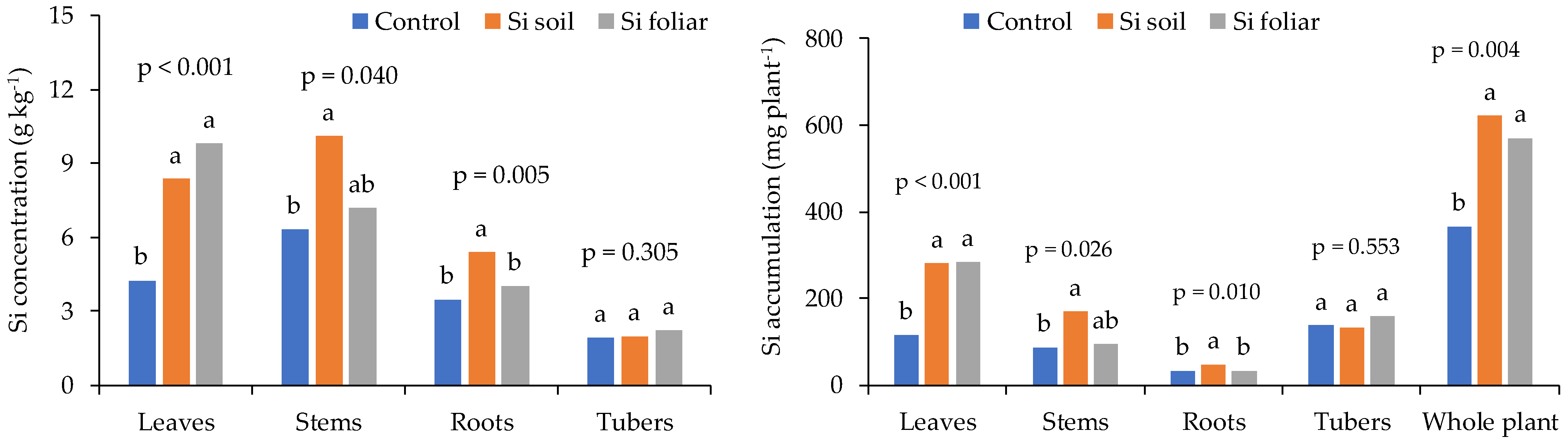 Figure 3. Comparison of soil- and foliar-applied sodium metasilicate on Si concentration and accumulation in potato leaves, roots, stems, and tubers [39]. Bars having common letters do not differ significantly at 5% level of significance.
In the same way, maximum uptake and accumulation of Si in cowpea (Vigna unguiculata) roots and leaves were observed for 800 g/kg soil-applied Si. Application of sodium metasilicate increased leaf Si concentration up to 4259.7 µg/g and root Si concentration up to 3126 µg/g [9].
Figure 3. Comparison of soil- and foliar-applied sodium metasilicate on Si concentration and accumulation in potato leaves, roots, stems, and tubers [39]. Bars having common letters do not differ significantly at 5% level of significance.
In the same way, maximum uptake and accumulation of Si in cowpea (Vigna unguiculata) roots and leaves were observed for 800 g/kg soil-applied Si. Application of sodium metasilicate increased leaf Si concentration up to 4259.7 µg/g and root Si concentration up to 3126 µg/g [9].
| Plant Species | Si% in Plant Biomass |
|---|---|
| Rice (O. sativa) | 4.17 |
| Wheat (Triticum aestivum) | 2.45 |
| Barley (Hordeum vulgare) | 1.82 |
| Tomato (Lycopersicon esculentum) | 1.54 |
| Sugarcane (Saccharum officianum) | 1.51 |
| Soybean (Glycine max) | 1.39 |
| Lettuce (Lactuca serriola) | 0.97 |
| Corn (Zea mays) | 0.82 |
| Potato (Solanum tuberosum) | 0.4 |
Figure 3. Comparison of soil- and foliar-applied sodium metasilicate on Si concentration and accumulation in potato leaves, roots, stems, and tubers [39]. Bars having common letters do not differ significantly at 5% level of significance.
References
- Guntzer, F.; Keller, C.; Meunier, J.D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213.
- Ma, J.F.; Takahashi, E. Soil, Fertilizer, and Plant Silicon Research in Japan; Elsevier: Amsterdam, The Netherlands, 2002.
- Tubana, B.S.; Babu, T.; Datnoff, L.E. A review of silicon in soils and plants and its role in us agriculture: History and future perspectives. Soil Sci. 2016, 181, 393–411.
- Sandhya, K.; Prakash, N.B.; Meunier, J.D.; Sandhya, K.; Prakash, N.B.; Meunier, J.D. Diatomaceous earth as source of silicon on the growth and yield of rice in contrasted soils of Southern India. J. Soil Sci. Plant Nutr. 2018, 18, 344–360.
- Artyszak, A. Effect of Silicon Fertilization on Crop Yield Quantity and Quality—A Literature Review in Europe. Plants 2018, 7, 54.
- Janislampi, K.W. Effect of Silicon on Plant Growth and Droucht Stress Tolerance. All Graduate Theses and Dissertations, UtahState University, Logan, UT, USA, 2012.
- Amin, M.; Ahmad, R.; Ali, A.; Hussain, I.; Mahmood, R.; Aslam, M.; Lee, D.J. Influence of Silicon Fertilization on Maize Performance Under Limited Water Supply. Silicon 2018, 10, 177–183.
- Cuong, T.X.; Ullah, H.; Datta, A.; Hanh, T.C. Effects of Silicon-Based Fertilizer on Growth, Yield and Nutrient Uptake of Rice in Tropical Zone of Vietnam. Rice Sci. 2017, 24, 283–290.
- Mali, M.; Aery, N.C. Effect of silicon on growth, biochemical constituents, and mineral nutrition of cowpea. Commun. Soil Sci. Plant Anal. 2009, 40, 1041–1052.
- Mehrabanjoubani, P.; Abdolzadeh, A.; Sadeghipour, H.R.; Aghdasi, M. Impacts of silicon nutrition on growth and nutrient status of rice plants grown under varying zinc regimes. Theor. Exp. Plant Physiol. 2015, 27, 19–29.
- Neu, S.; Schaller, J.; Dudel, E.G. Silicon availability modifies nutrient use efficiency and content, C:N:P stoichiometry, and productivity of winter wheat (Triticum aestivum L.). Sci. Rep. 2017, 7, 3–10.
- Luo, W.; Ma, J.; Khan, M.A.; Liao, S.; Ruan, Z.; Liu, H.; Zhong, B.; Zhu, Y.; Duan, L.; Fu, L.; et al. Cadmium accumulation in rice and its bioavailability in paddy soil with application of silicon fertilizer under different water management regimes. Soil Use Manag. 2021, 37, 299–306.
- Cai, Y.; Zhang, S.; Cai, K.; Huang, F.; Pan, B.; Wang, W. Cd accumulation, biomass and yield of rice are varied with silicon application at different growth phases under high concentration cadmium-contaminated soil. Chemosphere 2020, 242, 125128.
- Wang, B.; Chu, C.; Wei, H.; Zhang, L.; Ahmad, Z.; Wu, S.; Xie, B. Ameliorative effects of silicon fertilizer on soil bacterial community and pakchoi (Brassica chinensis L.) grown on soil contaminated with multiple heavy metals. Environ. Pollut. 2020, 267, 115411.
- Zhu, Y.; Gong, H. Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sustain. Dev. 2014, 34, 455–472.
- Santi, L.P.; Nurhaimi-Haris; Mulyanto, D. Effect of bio-silicon on drought tolerance in plants. In Proceedings of the Earth and Environmental Science, IOP Conference Series: Earth and Environmental Science, Jakarta, Indonesia, 18–20 October 2017; IOP Publishing: Bristol, UK, 2018; 183, p. 12014.
- Ahmad, M.; El-Saeid, M.H.; Akram, M.A.; Ahmad, H.R.; Haroon, H.; Hussain, A. Silicon fertilization—A tool to boost up drought tolerance in wheat (Triticum aestivum L.) crop for better yield. J. Plant Nutr. 2016, 39, 1283–1291.
- Shakoor, S.A.; Bhat, M.A.; Mir, S.H. Phytoliths in Plants: A Review. J. Bot. Sci. 2014, 3, 10–24.
- Green, S.W.; Piperno, D.R. Phytolith Analysis: An Archaeological and Geological Perspective. Am. J. Archaeol. 1991, 95, 741.
- Keeping, M.G. Uptake of Silicon by Sugarcane from Applied Sources May Not Reflect Plant-Available Soil Silicon and Total Silicon Content of Sources. Front. Plant Sci. 2017, 8, 1–14.
- Khan, M.A.; Goyal, V.; Jain, N. Impact of Ortho Silicic Acid Formulation on Yield and Disease incidence of Potatoes. In Proceedings of the 7th International Conference on Silicon in Agriculture, Bengaluru, India, 24–28 October 2017; p. 137.
- Kaya, C.; Tuna, L.; Higgs, D. Effect of silicon on plant growth and mineral nutrition of maize grown under water-stress conditions. J. Plant Nutr. 2006, 29, 1469–1480.
- Korndörfer, G.H.; Lepsch, I. Effect of silicon on plant growth and crop yield. Stud. Plant Sci. 2001, 8, 133–147.
- Ashtiani, F.A.; Kadir, J.; Nasehi, A.; Rahaghi, S.R.H.; Sajili, H. Effect of silicon on rice blast disease. Pertanika J. Trop. Agric. Sci. 2012, 35, 1–12.
- Ning, D.; Song, A.; Fan, F.; Li, Z.; Liang, Y. Effects of slag-based silicon fertilizer on rice growth and brown-spot resistance. PLoS ONE 2014, 9, e102681.
- Rodrigues, F.A.; Datnoff, L.E. Silicon and Plant Diseases; Springer: Berlin/Heidelberg, Germany, 2015; ISBN 9783319229300.
- Ahammed, G.J.; Yang, Y. Mechanisms of silicon-induced fungal disease resistance in plants. Plant Physiol. Biochem. 2021, 165, 200–206.
- Coskun, D.; Deshmukh, R.; Sonah, H.; Menzies, J.G.; Reynolds, O.; Ma, J.F.; Kronzucker, H.J.; Bélanger, R.R. The controversies of silicon’s role in plant biology. New Phytol. 2019, 221, 67–85.
- Hodson, M.J.; White, P.J.; Mead, A.; Broadley, M.R. Phylogenetic Variation in the Silicon Composition of Plants. Ann. Bot. 2005, 96, 1027–1046.
- Mandlik, R.; Thakral, V.; Raturi, G.; Shinde, S.; Nikolić, M.; Tripathi, D.K.; Sonah, H.; Deshmukh, R. Significance of Silicon Uptake, Transport, and Deposition in plants. J. Exp. Bot. 2020, 71, 6703–6718.
- Makabe, S.; Kakuda, K.; Sasaki, Y.; Ando, T.; Fujii, H.; Ando, H. Relationship between mineral composition or soil texture and available silicon in alluvial paddy soils on the Shounai Plain, Japan. Soil Sci. Plant Nutr. 2009, 55, 300–308.
- Agostinho, F.; Tubana, B.; Martins, M.; Datnoff, L. Effect of Different Silicon Sources on Yield and Silicon Uptake of Rice Grown under Varying Phosphorus Rates. Plants 2017, 6, 35.
- Wang, M.; Wang, R.; Mur, L.A.J.; Ruan, J.; Shen, Q.; Guo, S. Functions of silicon in plant drought stress responses. Hortic. Res. 2021, 8, 1–13.
- Liu, X.; Yin, L.; Deng, X.; Gong, D.; Du, S.; Wang, S.; Zhang, Z. Combined application of silicon and nitric oxide jointly alleviated cadmium accumulation and toxicity in maize. J. Hazard. Mater. 2020, 395, 122679.
- Zajaczkowska, A.; Korzeniowska, J.; Sienkiewicz-Cholewa, U. Effect of soil and foliar silicon application on the reduction of zinc toxicity in wheat. Agriculture 2020, 10, 522.
- Ranjan, A.; Sinha, R.; Bala, M.; Pareek, A.; Singla-Pareek, S.L.; KumarSingh, A. Silicon-mediated abiotic and biotic stress mitigation in plants: Underlying mechanisms and potential for stress resilient agriculture. Plant Physiol. Biochem. 2021, 163, 15–25.
- Debona, D.; Rodrigues, F.A.; Datnoff, L.E. Silicon’s Role in Abiotic and Biotic Plant Stresses. Annu. Rev. Phytopathol. 2017, 55, 85–107.
- Gong, H.; Chen, K. The regulatory role of silicon on water relations, photosynthetic gas exchange, and carboxylation activities of wheat leaves in field drought conditions. Acta Physiol. Plant. 2012, 34, 1589–1594.
- Pilon, C.; Soratto, R.P.; Moreno, L.A. Effects of soil and foliar application of soluble silicon on mineral nutrition, gas exchange, and growth of potato plants. Crop Sci. 2013, 53, 1605–1614.
- Zaheer, M.M.; Yasin, N.A.; Ahmad, S.R.; Khan, W.U.; Ahmad, A.; Ali, A.; Rehman, S.U. Amelioration of cadmium stress in gladiolus (Gladiolus grandiflora L.) by application of potassium and silicon. J. Plant Nutr. 2018, 41, 461–476.
- Bocharnikova, E.A.; Khomiakov, D.M.; Zhang, P.; Matichenkov, V.V.; Liu, Y.; Pakhnenko, E.P. Effect of Amorphous Silicon Dioxide on Cadmium Behavior in the Soil–Rice Plant System. Moscow Univ. Soil Sci. Bull. 2018, 73, 34–38.
- Greger, M.; Landberg, T.; Vaculík, M. Silicon Influences Soil Availability and Accumulation of Mineral Nutrients in Various Plant Species. Plants 2018, 7, 41.
- Savant, N.K.; Datnoff, L.E.; Snyder, G.H. Depletion of plant-available silicon in soils: A possible cause of declining rice yields. Commun. Soil Sci. Plant Anal. 1997, 28, 1245–1252.
- Taha, R.S.; Seleiman, M.F.; Shami, A.; Alhammad, B.A.; Mahdi, A.H.A. Integrated Application of Selenium and Silicon Enhances Growth and Anatomical Structure, Antioxidant Defense System and Yield of Wheat Grown in Salt-Stressed Soil. Plants 2021, 10, 1040.
- Savant, N.K.; Snyder, G.H.; Datnoff, L.E. Silicon Management and Sustainable Rice Production. Adv. Agron. 1997, 58, 151–199.
- Maghsoudi, K.; Emam, Y.; Ashraf, M. Foliar Application of Silicon at Different Growth Stages Alters Growth and Yield of Selected Wheat. J. Plant Nutr. 2016, 39, 1194–1203.
- Sirisuntornlak, N.; Ullah, H.; Sonjaroon, W.; Anusontpornperm, S.; Arirob, W.; Datta, A. Interactive Effects of Silicon and Soil pH on Growth, Yield and Nutrient Uptake of Maize. Silicon 2021, 13, 289–299.
- Walsh, O.S.; Shafian, S.; Mcclintick-chess, J.R.; Belmont, K.M.; Blanscet, S.M. Potential of Silicon Amendment for Improved Wheat Production. Plants 2018, 7, 26.
- Owino-Gerroh, C.; Gascho, G.J. Effect of Silicon on Low pH Soil Phosphorus Sorption and on Uptake and Growth of Maize. Commun. Soil Sci. Plant Anal. 2005, 35, 2369–2378.
- Pavlovic, J.; Kostic, L.; Bosnic, P.; Kirkby, E.A.; Nikolic, M. Interactions of Silicon with Essential and Beneficial Elements in Plants. Front. Plant Sci. 2021, 12, 697592.
- Gou, T.; Yang, L.; Hu, W.; Chen, X.; Zhu, Y.; Guo, J.; Gong, H. Silicon improves the growth of cucumber under excess nitrate stress by enhancing nitrogen assimilation and chlorophyll synthesis. Plant Physiol. Biochem. 2020, 152, 5361.
- Hasanuzzaman, M.; Fujita, M.; Oku, H.; Nahar, K.; Hawrylak-Nowak, B. Nutrients and Abiotic Stress Tolerance; Springer: Singapore, 2018; ISBN 9789811090431.
- Pati, S.; Pal, B.; Badole, S.; Hazra, G.C. Effect of Silicon Fertilization on Growth, Yield, and Nutrient Uptake of Rice. Commun. Soil Sci. Plant Anal. 2016, 3624, 1532–2416.
- Eneji, A.E.; Inanaga, S.; Muranaka, S.; Li, J.; Hattori, T.; An, P.; Tsuji, W.; Eneji, A.E.; Inanaga, S.; Muranaka, S.; et al. Growth and Nutrient Use in Four Grasses Under Drought Stress as Mediated by Silicon Fertilizers. J. Plant Nutr. 2008, 31, 355–365.
- Pulz, A.L.; Crusciol, C.A.C.; Lemos, L.B.; Soratto, R.P. Silicate and Limestone Effects on Potato Nutrition, Yield and Quality under Drought Stress. Rev. Bras. Ciência do Solo 2008, 32, 1651–1659.
- Reboredo, F.; Lidon, F.C.; Pessoa, F.; Duarte, M.P.; Silva, M.J. The uptake of macronutrients by an active silicon accumulator plant growing in two different substrata. Emirates J. Food Agric. 2013, 25, 986–993.
- Ma, J.F. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr. 2004, 50, 11–18.
- Gouharrizi, Z.Z.; Khorassani, R.; Halajnia, A. The effect of silicon on phosphorus uptake and wheat growth under drought stress in a calcareous soil. Environ. Stresses Crop Sci. 2021, 14(3), 665–673.
- Ma, J.F.; Takahashi, E. Function of silicon in plant growth. In Soil, Fertilizer and Plant Silicon Research in Japan, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2002; pp. 151–154.
- Hawk, C.; Hyland, J.; Rupert, R.; Colonvega, M.; Hall, S. Assessment of balance and risk for falls in a sample of community-dwelling adults aged 65 and older. Chiropr. Osteopat. 2006, 14, 3.
- Vieira da Cunha, K.P.; Williams Araújo do Nascimento, C.; José da Silva, A. Silicon alleviates the toxicity of cadmium and zinc for maize (Zea mays L.) grown on a contaminated soil. J. Plant Nutr. Soil Sci. 2008, 171, 849–853.
- Fan, X.; Wen, X.; Huang, F.; Cai, Y.; Cai, K. Effects of silicon on morphology, ultrastructure and exudates of rice root under heavy metal stress. Acta Physiol. Plant. 2016, 38, 197.
- Li, J.; Leisner, M.; Frantz, J. Alleviation of copper toxicity in Arabidopsis thaliana by silicon addition to hydroponic solutions. J. Am. Soc. Hortic. Sci. 2008, 133, 70–77.
- Flora, C.; Khandekar, S.; Boldt, J.; Leisner, S. Silicon alleviates long-term copper toxicity and influences gene expression in Nicotiana tabacum. J. Plant Nutr. 2019, 42, 864–878.
- Kim, Y.H.; Khan, A.L.; Kim, D.H.; Lee, S.Y.; Kim, K.M.; Waqas, M.; Jung, H.Y.; Shin, J.H.; Kim, J.G.; Lee, I.J. Silicon mitigates heavy metal stress by regulating P-type heavy metal ATPases, Oryza sativa low silicon genes, and endogenous phytohormones. BMC Plant. Boil. 2014, 14, 13.
- Savić, J.; Marjanović-Jeromela, A. Effect of silicon on sunflower growth and nutrient accumulation under low boron supply. Helia 2013, 36, 61–68.
- Elsokkary, I.H. Silicon as a Beneficial Element and as an Essential Plant Nutrient: An Outlook (Review). Alexandria Sci. Exch. J. 2018, 39, 534–550.
- Rastogi, A.; Tripathi, D.K.; Yadav, S.; Chauhan, D.K.; Živčák, M.; Ghorbanpour, M.; El-Sheery, N.I.; Brestic, M. Application of silicon nanoparticles in agriculture. 3 Biotech. 2019, 9, 90.
More