Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Camila Xu and Version 1 by Workie Anley Zegeye.
CRISPR/Cas9 is an endonuclease of DNA that splits the invading phage DNA into pieces and then incorporates it into the CRISPR set as a spacer. It was implemented efficiently in plants in 2013, and in five original research articles, the CRISPR/Cas9 scheme in rice was efficiently recorded.
- CRISPR/Cas9
- Cas variants
- genome editing
1. CRISPR/Cas9 System and Its Components
Initially, CRISPR/Cas9 was discovered as an adaptive immune system of archaea and bacteria. It was first reported by [31][1], and researchers discovered a means of exploiting it as a gene-editing technology. This technique can identify a particular site in a target gene in a highly efficient, unique, and flexible manner [32][2]. In roughly 50% of bacteria and 90% of archaea, CRISPR/Cas9 active immune mechanisms exist [33][3]. CRISPR/Cas9 is an endonuclease of DNA that splits the invading phage DNA into pieces and then incorporates it into the CRISPR set as a spacer. It was implemented efficiently in plants in 2013, and in five original research articles, the CRISPR/Cas9 scheme in rice was efficiently recorded [4,19,34,35,36][4][5][6][7][8]. Consequently, CRISPR/Cas9 technologies have proved to be an essential genome-editing method for rice.
Nowadays, the editing system of CRISPR/Cas9 is the most common mechanism in plant biology for the genome editing process [37,38][9][10] because of its uncomplicatedness, flexibility, and efficiency. Two CRISPR/Cas9 components: the Cas9 protein and a short RNA molecule (sgRNA), together form a ribonucleoprotein complex (Figure 1). The sgRNA is made up of 18 to 21 nucleotides that are designed to target specific sites in the genome (protospacer). A G-rich (5′-NGG-3′) protospacer-adjacent motif (PAM) should be found downstream of the target site [31][1]. tracrRNA (trans-activating CRISPR RNA) and crRNA (CRISPR-derived RNA) are the two principal components of guide RNA. In essence, crRNA has homology area that enables it to integrate with tracrRNA. The tracrRNA has a stem-loop shape associated with the Cas9 protein. The crRNA and tracrRNA should be designed as sgRNA in the CRISPR/Cas9 gene-editing scheme to direct Cas9 dsDNA cleavage at the targeted site [31][1]. Following the appropriate site identification by sgRNA based on the Watson–Crick base-pairing rule, the Cas9-sgRNA complex moves through the genome and produces a double-stranded break (DSB) [33][3]. During cleavage site repairing, the error-prone non-homologous end-joining (NHEJ) pathway regularly leaves a lesion in the form of a minor InDel or substitution upstream of PAM. Such mutations can induce frame-shift mutations in the coding sequence of the gene and cause a premature stop codon, resulting in a loss or gain of function in mutants. This achievement has unlocked numerous other possibilities for scientists to obtain more knowledge on plant biological systems [39][11].
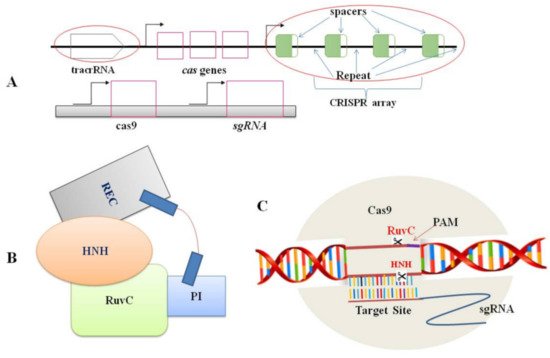
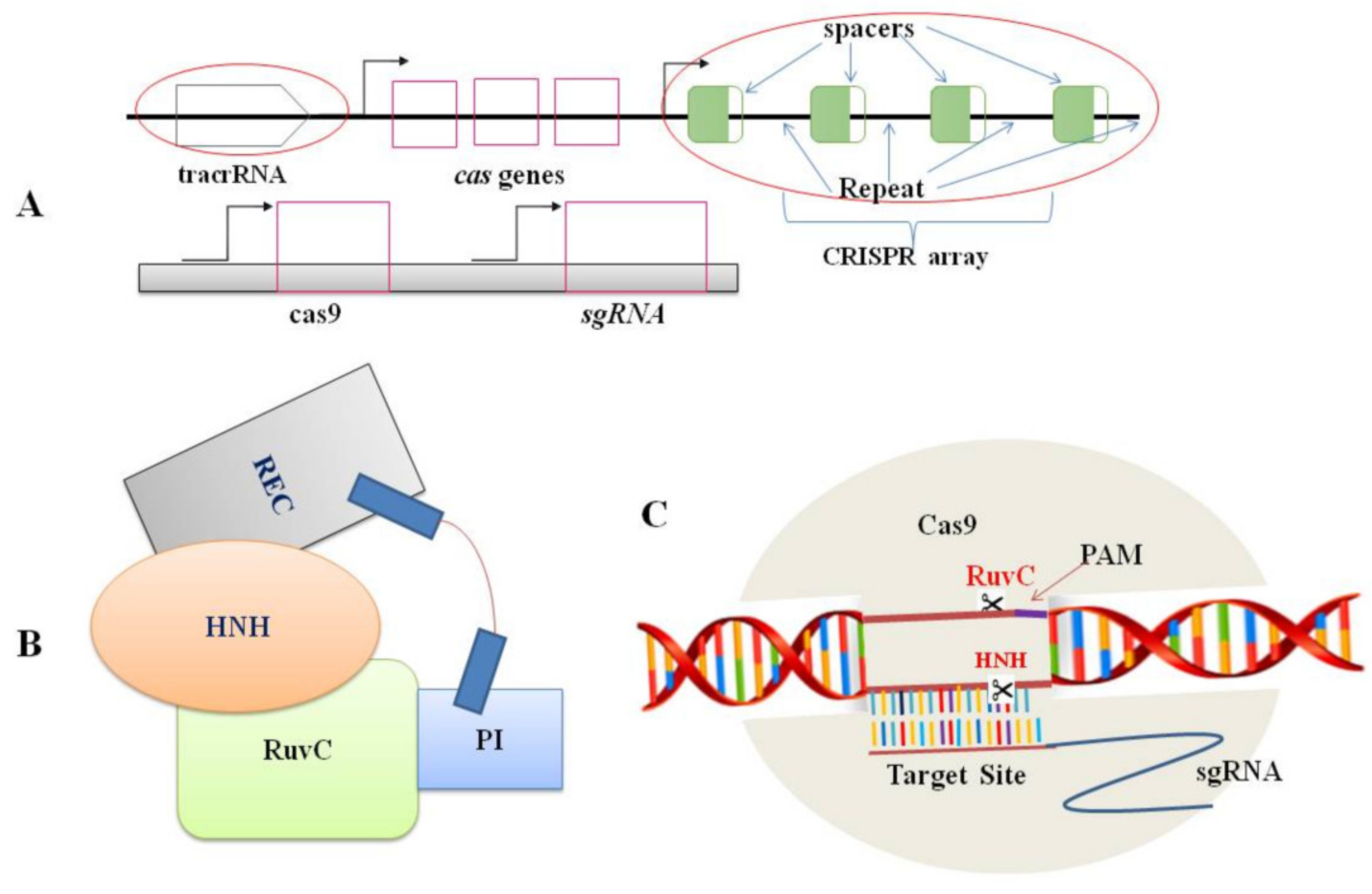 Figure 1. Components of CRISPR/Cas9 system: (A). Genomic structures of the CRISPR/Cas system (top) and the engineered CRISPR/Cas9 system (bottom); (B). A schematic representation of the Cas9 protein structure. Domains includes REC (large recognition lobe) and RuvC (a nuclease domain), which is linked with an arginine-rich region. HNH is a second nuclease domain. PI is PAM-interacting domain; (C). The conformation of the Cas9–sgRNA complex in the process of DNA cleavage. The Cas9 endonuclease is targeted to DNA by a guide RNA which can be supplied as a two-part system consisting of crRNA and tracrRNA or as a single guide RNA, where the crRNA and tracrRNA are connected by a linker. Target recognition is facilitated by the protospacer-adjacent motif (PAM). Cleavage occurs on both strands (scissors) 3 bp upstream of the PAM.
Figure 1. Components of CRISPR/Cas9 system: (A). Genomic structures of the CRISPR/Cas system (top) and the engineered CRISPR/Cas9 system (bottom); (B). A schematic representation of the Cas9 protein structure. Domains includes REC (large recognition lobe) and RuvC (a nuclease domain), which is linked with an arginine-rich region. HNH is a second nuclease domain. PI is PAM-interacting domain; (C). The conformation of the Cas9–sgRNA complex in the process of DNA cleavage. The Cas9 endonuclease is targeted to DNA by a guide RNA which can be supplied as a two-part system consisting of crRNA and tracrRNA or as a single guide RNA, where the crRNA and tracrRNA are connected by a linker. Target recognition is facilitated by the protospacer-adjacent motif (PAM). Cleavage occurs on both strands (scissors) 3 bp upstream of the PAM.
2. Mechanism of the CRISPR/Cas9 System
In three steps, namely adaptation/acquisition, expression, and interference, CRISPR/Cas9 identifies and targets the genetic material of foreign DNA [40,41,42,43][12][13][14][15]. The adaptation/acquisition phase includes the recognition, invasion, and binding of donor DNA that are cut into small segments and combined within the CRISPR locus. Then, the CRISPR locus is transcribed to create crRNA that directs the intended effect or endonucleases to attack viral items by complementary base pairing [44,45][16][17]. Since the protospacer contains a G-rich base pair (5′-NGG-3′), PAMs are used as recognition motifs for the adaptation/acquisition of the targeted site. During the second step of the CRISPR/Cas9 execution mechanism, the lengthy Pre-crRNA is deliberately transcribed from the CRISPR nucleus and reproduced into crRNAs using Cas9 proteins. Recently, researchers revealed that tracrRNA is also involved in Streptococcus pyogenes pre-crRNA processing [46][18]. The tracrRNA is associated with the repeat crRNA site through complementary base pairing and allows precrRNA to be processed in crRNA [47][19]. The processed crRNAs join the associated antiviral defense complex of CRISPR and help to identify and parse a particular target area of donor DNA [47][19]. At the final steps, i.e., interference, the system needs a Cas9 protein [48,49][20][21] so that the sgRNA can guide the cleavage of the Cas9 protein complex from the particular target area; this creates immunity from pathogen attacks [40,41][12][13].
3. The Advantages and Drawbacks of the CRISPR/Cas9 System
Until recently, the CRISPR/Cas9 tool was thought to be the best option for genome editing (GE) in plants, but it still has some drawbacks that limit its widespread application (Figure 2).
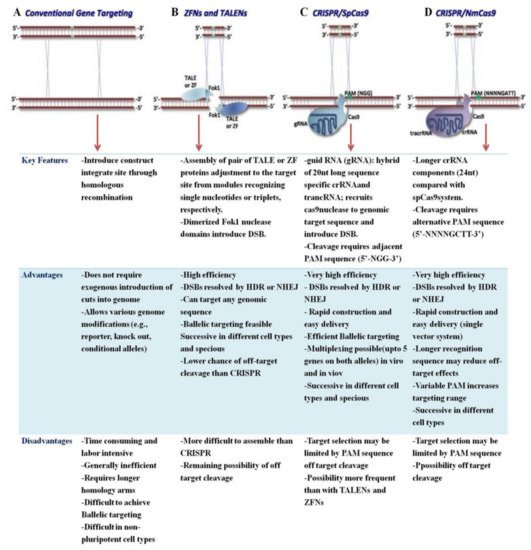
 Figure 2. The advantages and disadvantages of the CRISPR/Cas9 system over other approaches for genome editing. (A). Conventional gene targeting. (B). ZNFs and TALENs. (C). CRISPR/SpCas9. (D). CRISPR/NmCas9. The red arrow indicates the corresponding gene editing method with its features, advantages, and disadvantages.
Figure 2. The advantages and disadvantages of the CRISPR/Cas9 system over other approaches for genome editing. (A). Conventional gene targeting. (B). ZNFs and TALENs. (C). CRISPR/SpCas9. (D). CRISPR/NmCas9. The red arrow indicates the corresponding gene editing method with its features, advantages, and disadvantages.
The following are a list of the major issues with CRISPR/Cas9 and the advantages of CRISPR/Cas variants:
-
The CRISPR/Cas9 system’s large size limits its editing efficiency, and it is not suitable for packing into viral vectors for delivery to somatic tissues. For efficient plant GE, a smaller-sized CRISPR/Cas is required;
-
SpCas9 involves a 5′-NGG-3′ PAM next to a 20 nt DNA target site where it only distinguishes the NGG PAM sequence, which limits its effectiveness when compared to new CRISPR/Cas variants. NG-Cas9 is more active and there is a newer variant, SpRYCas9 that is almost PAM-less. The broad PAM compatibility of SpRY greatly expands the targeting scope of CRISPR-based tools in plant genome engineering;
-
CRISPR/Cas9 has the potential to incorporate a large number of off-target mutations into the genome. However, by identifying various PAMs, new CRISPR/Cas variants have achieved better editing efficiency (fewer off-target mutations) of target bases in the sequence of interest;
-
CRISPR/Cas9 generates mutations at non-specific loci that are homologous to target sites;
-
CRISPR/Cas9-made mutant plants via Agrobacterium-mediated transformation systems are more expensive, time consuming, and resource intensive. The use of tissue culture-free genome editing systems, on the other hand, has the potential to improve efficiency;
-
The commercialization of transgenic crops expressing CRISPR/Cas9 faces challenges in a number of countries, owing primarily to development costs and constraints imposed by regulatory systems for the field release of genetically modified organisms.