Ramularia leaf spot (RLS), caused by the fungus Ramularia collo-cygni, has recently become widespread in Europe. Succinate dehydrogenase inhibitor (SDHI) and demethylation inhibitor (DMI) fungicides are mainly applied for disease control on barley fields, but pathogen isolates with a reduced sensitivity can cause difficulties. There is an urgent need for new spring barley cultivars that are more resistant to RLS development and can inhibit R. collo-cygni epidemics.
- Ramularia leaf spot
- fungicide target proteins
- CYP51
- azoles
- SDHI
- mlo gene
1. Introduction
Barley (Hordeum vulgare L.) is one of the most important cereal crops grown in temperate regions worldwide
. Among the various pests and diseases that threaten sustainable barley cultivation
, Ramularia leaf spot (RLS)—caused by an ascomycete, Ramularia collo-cygni—has become a new threat to barley cultivation. The first reported case of the disease dates back to 1893 in northern Italy and was identified by a notable botanist, Fridiano Cavara. For a century, RLS was a minor disease and did not cause any serious problems in barley cultivation, but the majority of R. collo-cygni outbreaks have been reported in the last few decades. The first official records in Germany, the UK, Ireland, and New Zealand are from the 1990s. In Finland, Sweden, Denmark, and France, the first records are from the 2000s, and in Estonia, Spain, and Australia, the first records are after 2010
. Since the beginning of 2000, RLS has been considered to be an emerging disease of barley in Europe, South America, and New Zealand
2. Epidemiology of Ramularia collo-cygni
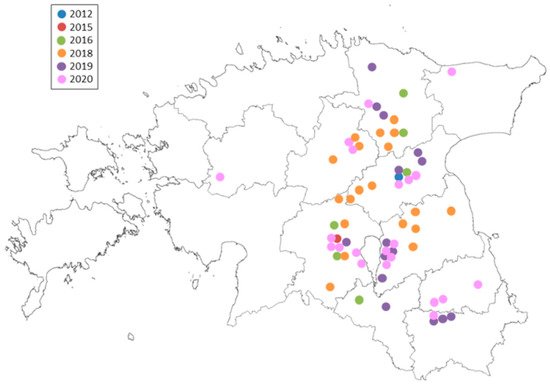
3. Fungicide Resistance in R. collo-cygni Populations
3.1. Status of DMI-Fungicide Sensitivity
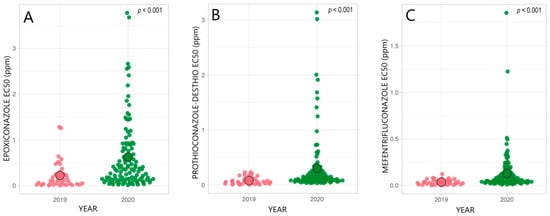
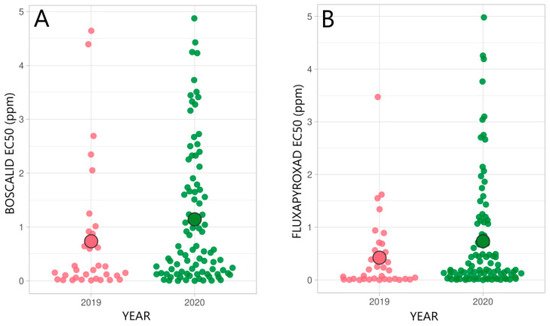
3.2. Status of SDHI-Fungicide Sensitivity
. It is possible that RLS can lead to moderate barley yield losses of 5–20% or greater. Although the trigger of these increasing RLS epidemics is still under debate, R. tritici
4. Common Barley Cultivars Enhance RLS Epidemics
5. Pleiotropic Effect of mlo Alleles on RLS
adaptation to widely distributed RLS-susceptible barley cultivars and fungicides along with heat stress under global climate change will lead to a risk of future RLS epidemics
6. Future Challenges
.
References
- Newton, A.C.; Flavell, A.J.; George, T.; Leat, P.; Mullholland, B.; Ramsay, L.; Revoredo-Giha, C.; Russell, J.; Steffenson, B.J.; Swanston, J.S.; et al. Crops that feed the world 4. Barley: A resilient crop? Strengths and weaknesses in the context of food security. Food Secur. 2011, 3, 141–178.
- Walters, D.R.; Havis, N.D.; Oxley, S.J. Ramularia collo-cygni: The biology of an emerging pathogen of barley. FEMS Microbiol. Lett. 2008, 279, 1–7.
- Dussart, F.; Creissen, H.E.; Havis, N.D. Ramularia collo-cygni—An Enemy in Waiting. In eLS; 2020; pp. 1–8. Available online: https://pure.sruc.ac.uk/en/publications/riamularia-collo-cygnii-an-enemy-in-waiting (accessed on 13 October 2021).
- Havis, N.; Brown, J.K.M.; Clemente, G.; Frei, P.; Jedryczka, M.; Kaczmarek, J.; Kaczmarek, M.; Matusinsky, P.; McGrann, G.; Pereyra, S.; et al. Ramularia collo-cygni—An Emerging Pathogen of Barley Crops. Phytopathology 2015, 105, 895–904.
- McGrann, G.D.; Havis, N. Ramularia Leaf Spot: A Newly Important Threat to Barley Production. Outlooks Pest Manag. 2017, 28, 65–69.
- West, J.S.; Townsend, J.A.; Stevens, M.; Fitt, B.D.L. Comparative biology of different plant pathogens to estimate effects of climate change on crop diseases in Europe. Eur. J. Plant Pathol. 2012, 133, 315–331.
- Matusinsky, P.; Leisova-Svobodova, L.; Gubis, J.; Hudcovicova, M.; Klcova, L.; Gubisova, M.; Marik, P.; Tvaruzek, L.; Minarikova, V. Impact of the seed-borne stage of Ramularia collo-cygni in barley seed. J. Plant Pathol. 2011, 93, 679–689.
- Havis, N.D.; Oxley, S.J.P. Investigation the life cycle of Ramularia collo-cygni using PCR diagnostics. In First European Ramularia Workshop 12–14 March 2006; Koopmann, B., Oxley, S., Schützendübel, A., von Tiedemann, A., Eds.; Georg-August-Universität: Göttingen, Germany, 2006; pp. 39–44.
- Kaczmarek, M.; Piotrowska, M.; Fountaine, J.; Gorniak, K.; McGrann, G.; Armstrong, A.; Wright, K.; Newton, A.; Havis, N. Infection strategy of Ramularia collo-cygni and development of Ramularia leaf spot on barley and alternative graminaceous hosts. Plant Pathol. 2016, 66, 45–55.
- Stabentheiner, E.; Minihofer, T.; Huss, H. Infection of Barley by Ramularia collo-cygni: Scanning Electron Microscopic Investigations. Mycopathologia 2009, 168, 135–143.
- Havis, N.D.; Nyman, M.; Oxley, S.J.P. Evidence for seed transmission and symptomless growth of R amularia collo-cygni in barley (H ordeum vulgare). Plant Pathol. 2013, 63, 929–936.
- Rodriguez, R.; Redman, R. More than 400 million years of evolution and some plants still can’t make it on their own: Plant stress tolerance via fungal symbiosis. J. Exp. Bot. 2008, 59, 1109–1114.
- Schützendübel, A.; Stadler, M.; Wallner, D.; von Tiedemann, A. A hypothesis on physiological alterations during plant ontogenesis governing susceptibility of winter barley to Ramularia leaf spot. Plant Pathol. 2008, 57, 518–526.
- Harvey, I. Epidemiology and control of leaf and awn spot of barley caused by Ramularia collocygni. N. Z. Plant Prot. 2002, 55, 331–335.
- Sjökvist, E.; Lemcke, R.; Kamble, M.; Turner, F.; Blaxter, M.; Havis, N.; Lyngkjær, M.; Radutoiu, S. Dissection of Ramularia Leaf Spot Disease by Integrated Analysis of Barley and Ramularia collo-cygni Transcriptome Responses. Mol. Plant-Microbe Interact. 2019, 32, 176–193.
- Brown, J.K.M.; Rant, J.C. Fitness costs and trade-offs of disease resistance and their consequences for breeding arable crops. Plant Pathol. 2013, 62, 83–95.
- McGrann, G.; Stavrinides, A.; Russell, J.; Corbitt, M.M.; Booth, A.; Chartrain, L.; Thomas, W.T.B.; Brown, J.K.M. A trade off between mlo resistance to powdery mildew and increased susceptibility of barley to a newly important disease, Ramularia leaf spot. J. Exp. Bot. 2014, 65, 1025–1037.
- Leistrumaitė, A.; Liatukas, Ž. Resistance of spring barley cultivars to the new disease Ramularia leaf spot, caused by Ramularia collo-cygni. Agron. Res. 2006, 4, 251–255.
- Sooväli, P.; Tikhonova, M.; Matušinsky, P. First Report of Ramularia Leaf Spot Caused by Ramularia collo-cygni on Leaves and Seeds of Barley in Estonia. Plant Dis. 2014, 98, 997.
- Kiiker, R.; Juurik, M.; Mäe, A. Fungicide Resistance Evolving in Ramularia collo-cygni Population in Estonia. Microorganisms 2021, 9, 1514.
- Leisova-Svobodova, L.; Matusinsky, P.; Kucera, L. Variability of the Ramularia collo-cygni Population in Central Europe. J. Phytopathol. 2012, 160, 701–709.
- Piotrowska, M.; Ennos, R.; Fountaine, J.; Burnett, F.; Kaczmarek, M.; Hoebe, P. Development and use of microsatellite markers to study diversity, reproduction and population genetic structure of the cereal pathogen Ramularia collo-cygni. Fungal Genet. Biol. 2016, 87, 64–71.
- Strobel, D.; Bryson, R.; Stammler, G.; Prochnow, J. A European overview of the occurrence of Ramularia collo-cygni and its sensitivity to fluxapyroxad. In Proceedings of the 11th Conference of the European Foundation for Plant Pathology–Healthy People, Krakow, Poland, 8–13 September 2014.
- Jørgensen, L.N.; Heick, T.M. Azole Use in Agriculture, Horticulture, and Wood Preservation–Is It Indispensable? Front. Cell. Infect. Microbiol. 2021, 11, 806.
- Stam, R.; Sghyer, H.; Tellier, A.; Hess, M.; Hückelhoven, R. The Current Epidemic of the Barley Pathogen Ramularia collo-cygni Derives from a Population Expansion and Shows Global Admixture. Phytopathology 2019, 109, 2161–2168.
- Kuck, K.-H.; Russell, P.E. FRAC: Combined resistance risk assessment. Asp. Appl. Biol. 2006, 78, 3–10.
- Fountaine, J.M.; Fraaije, B.A. Development of QoI resistant alleles in populations of Ramularia collo-cygni. In Proceedings of the Aspects of Applied Biology 92, The 2nd European Ramularia Workshop–A New Disease and Challenge in Barley Production; Association of Applied Biologists, The Warwick Enterprise Park Wellesbourne: Warwick/Edinburgh, UK, 2009; pp. 123–126.
- Matusinsky, P.; Svobodova-Leisova, L.; Mariks, P.; Tvaruzek, L.; Stemberkova, L.; Hanusova, M.; Minarikova, V.; Vysohlidova, M.; Spitzer, T. Frequency of a mutant allele of cytochrome b conferring resistance to Ool fungicides in the Czech population of Ramularia collo-cygni. J. Plant Dis. Prot. 2010, 117, 248–252.
- Havis, N.D.; Gorniak, K.; Taylor, J.; Stanisz-Migal, M.; Burnett, F.J. Controlling Ramularia leaf spot in barley crops. In Proceedings of the Proceedings Crop Production in Northern Britain 2018, Dundee, UK, 27–28 February 2018; The Association for Crop Protection in Northern Britain: Dundee, UK, 2018; pp. 91–96.
- Dooley, H.; Shaw, M.W.; Spink, J.; Kildea, S. The effect of succinate dehydrogenase inhibitor/azole mixtures on selection of Zymoseptoria triticiisolates with reduced sensitivity. Pest Manag. Sci. 2015, 72, 1150–1159.
- Clark, W.S. Septoria tritici and azole performance. Asp. Appl. Biol. 2006, 78, 127–132.
- Lucas, J.A.; Hawkins, N.J.; Fraaije, B.A. The Evolution of Fungicide Resistance. In Advances in Applied Microbiology; Elsevier Ltd.: Amsterdam, The Netherlands, 2015; Volume 90, pp. 29–92.
- Huf, A.; Rehfus, A.; Lorenz, K.H.; Bryson, R.; Voegele, R.T.; Stammler, G. Proposal for a new nomenclature for CYP51haplotypes inZymoseptoria triticiand analysis of their distribution in Europe. Plant Pathol. 2018, 67, 1706–1712.
- Rehfus, A.; Matusinsky, P.; Strobel, D.; Bryson, R.; Stammler, G. Mutations in target genes of succinate dehydrogenase inhibitors and demethylation inhibitors in Ramularia collo-cygni in Europe. J. Plant Dis. Prot. 2019, 126, 447–459.
- FRAC—Fungicide Resistance Action Committee. Available online: http://www.frac.info/ (accessed on 6 June 2019).
- Heick, T.M.; Matzen, N.; Jørgensen, L.N. Reduced field efficacy and sensitivity of demethylation inhibitors in the Danish and Swedish Zymoseptoria tritici populations. Eur. J. Plant Pathol. 2020, 157, 625–636.
- Kiiker, R.; Juurik, M.; Heick, T.; Mäe, A. Changes in DMI, SDHI, and QoI Fungicide Sensitivity in the Estonian Zymoseptoria tritici Population between 2019 and 2020. Microorganisms 2021, 9, 814.
- Piotrowska, M.J.; Fountaine, J.M.; Ennos, R.A.; Kaczmarek, M.; Burnett, F. Characterisation of Ramularia collo-cygni laboratory mutants resistant to succinate dehydrogenase inhibitors. Pest Manag. Sci. 2016, 73, 1187–1196.
- Rehfus, A.; Miessner, S.; Achenbach, J.; Strobel, D.; Bryson, R.; Stammler, G. Emergence of succinate dehydrogenase inhibitor resistance of Pyrenophora teres in Europe. Pest Manag. Sci. 2016, 72, 1977–1988.
- Dooley, H.; Shaw, M.W.; Mehenni-Ciz, J.; Spink, J.; Kildea, S. Detection of Zymoseptoria tritici SDHI-insensitive field isolates carrying the SdhC -H152R and SdhD -R47W substitutions. Pest Manag. Sci. 2016, 72, 2203–2207.
- Rehfus, A.; Strobel, D.; Bryson, R.; Stammler, G. Mutations in sdh genes in field isolates of Zymoseptoria tritici and impact on the sensitivity to various succinate dehydrogenase inhibitors. Plant Pathol. 2017, 67, 175–180.
- Büschges, R.; Hollricher, K.; Panstruga, R.; Simons, G.; Wolter, M.; Frijters, A.; van Daelen, R.; van der Lee, T.; Diergaarde, P.; Groenendijk, J.; et al. The Barley Mlo Gene: A Novel Control Element of Plant Pathogen Resistance. Cell 1997, 88, 695–705.
- Jørgensen, I.H. Discovery, characterization and exploitation of Mlo powdery mildew resistance in barley. Euphytica 1992, 63, 141–152.
- Lyngkjær, M.; Newton, A.C.; Atzema, J.L.; Baker, S.J. The Barley mlo-gene: An important powdery mildew resistance source. Agronomie 2000, 20, 745–756.
- Dreiseitl, A. Specific Resistance of Barley to Powdery Mildew, Its Use and Beyond. A Concise Critical Review. Genes 2020, 11, 971.
- Kjær, B.; Jensen, H.P.; Jensen, J.; Jørgensen, J.H. Associations between three ml-o powdery mildew resistance genes and agronomic traits in barley. Euphytica 1990, 46, 185–193.
- McGrann, G.R.D.; Burt, C.; Nicholson, P.; Brown, J.K.M. Differential effects of lesion mimic mutants in barley on disease development by facultative pathogens. J. Exp. Bot. 2015, 66, 3417–3428.
- Oxley, S.J.P.; Havis, N.D. Development of Ramularia collo-cygni on spring barley and its impact on yield. In Proceedings of the Dundee Conference: Crop protection in Northern Britain 2004, Dundee, UK, 24–25 February 2004; Heilbronn, T., Ed.; pp. 147–152.
- Rostoks, N.; Schmierer, D.; Mudie, S.; Drader, T.; Brueggeman, R.; Caldwell, D.G.; Waugh, R.; Kleinhofs, A. Barley necrotic locus nec1 encodes the cyclic nucleotide-gated ion channel 4 homologous to the Arabidopsis HLM1. Mol. Genet. Genom. 2005, 275, 159–168.
- McGrann, G.R.D.; Brown, J.K.M. The role of reactive oxygen in the development of Ramularia leaf spot disease in barley seedlings. Ann. Bot. 2017, 121, 415–430.
- McGrann, G.R.D.; Miller, S.; Havis, N. The enhanced magnaporthe resistance 1 locus affects Ramularia leaf spot development in barley. Eur. J. Plant Pathol. 2019, 156, 123–132.
- McGrann, G.R.D.; Steed, A.; Burt, C.; Goddard, R.; Lachaux, C.; Bansal, A.; Corbitt, M.; Gorniak, K.; Nicholson, P.; Brown, J.K.M. Contribution of the drought tolerance-related Stress-responsive NAC 1 transcription factor to resistance of barley to R amularia leaf spot. Mol. Plant Pathol. 2014, 16, 201–209.
- Bosch, F.V.D.; Paveley, N.; Shaw, M.; Hobbelen, P.; Oliver, R.A. The dose rate debate: Does the risk of fungicide resistance increase or decrease with dose? Plant Pathol. 2011, 60, 597–606.