Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Catherine Yang and Version 1 by Shirong Guo.
Salinity is a ubiquitous stressor, depleting osmotic potential and affecting the tomato seedlings’ development and productivity. The RBOH transcription factors activated the hydrogen peroxide-mediated signalling pathway that induced the detoxification mechanisms in tomato seedlings. Consequently, the increased gene expression of antioxidant enzymes and the corresponding ratio of non-enzymatic antioxidants AsA-GSH suggested the modulation of antioxidants to survive the salt-induced oxidative stress. In addition, the endogenous ABA level was enhanced under salinity stress, indicating higher ABA biosynthesis and signalling gene expression.
- tomato
- ABA
- RBOH
- ROS
1. Introduction
Salt stress is a major abiotic stress that adversely affects crop growth, development, quality, and yield [1,2][1][2]. High salinity affects plant cellular function by influencing uptake and assimilation of mineral ions, enzyme activity and photosynthetic function [3,4,5][3][4][5]. The higher accumulation of Na+ ions induces ionic and osmotic stress in plants, which causes alteration in plants structural and functional stability [6,7][6][7]. High salt concentrations in plants can cause an imbalance between production and scavenging of reactive oxygen species (ROS) such as superoxide anion (O2•−), hydrogen peroxide (H2O2) and hydroxyl radicals (OH•), particularly in chloroplasts and mitochondria, and induce hyperosmotic stress that can lead to oxidative damage [8,9][8][9]. In addition, the excess of salts reduces growth, mainly by reducing cell expansion in root tips and younger leaves thereby promoting premature senescence or programmed cell death [10,11][10][11].
Plants have developed various physiological, biochemical, and molecular processes to regulate salt stress’s adverse effects. Usually, plants endure sub-optimal environments by adopting a set of mechanisms that collectively respond against the stressful environment either by acclimation, escape, or detoxification [7,12][7][12]. Among them, one such possible mechanism is the accumulation of osmoprotectants, such as amino acids (proline) and non-structural sugars to maintain the osmotic balance under the prevailing stress conditions [13,14][13][14]. Besides this, plants also have an antioxidative defense system that contains enzymatic antioxidants (Superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), ascorbate peroxidase (APX), Glutathione peroxidase (GPX), glutathione reductase (GR)) and non-enzymatic antioxidants (Ascorbate(AsA) and Glutathione (GSH)) that helps in scavenging the stress-induced reactive oxygen species (ROSs) [15,16][15][16].
ROS also acts as a multipurpose signal to initiate a series of subsequent defense signalling under various environmental stresses, including high salinity, drought, pathogenic infection and heat stress [17]. Particularly, the NADPH oxidase/respiratory burst oxidase homolog (RBOHs) transcription factors are essential in regulating ROS generation and subsequent signalling events for salinity adaptation. Previous investigations revealed that RBOHs are involved in different signalling pathways that regulate root hair growth, stomatal closure, pollen–stigma interactions, defense responses to pathogens, and acclimation to abiotic stresses [18,19,20][18][19][20]. Notably, tomato consists of eight RBOHs that are involved in stomatal movements through different phytohormones. For instance, RBOH1 gene is linked to Abscisic acid (ABA)-mediated salinity tolerance in tomatoes. Accumulation of ABA usually increases transcript levels of RBOH1 and other defense-related genes resulting in elevated apoplastic H2O2 collection, increasing the activity of NADPH oxidase and antioxidant enzymes in tomatoes [21]. In addition to their importance in biotic and abiotic responses, RBOH genes are also shown to play critical roles in plant growth and development associated with hormone signalling [22].
ABA is a primary plant hormone involved in salt stress response regulation, including stomatal closure, ion homeostasis, salt stress-responsive gene expression, and metabolic changes [23]. ABA functions as a central integrator that links and reprograms the complex developmental process under salt stress and activates adaptive signalling cascades in plants [24]. Under stress conditions, plants induce the production of ABA biosynthesis genes, such as Nine-Cis-Epoxycarotenoid Dioxygenases (NCEDs) and ABA Deficient (ABAs). The ABA receptors then perceive ABA Pyrabactin Resistance/Pyrabactin Resistance Like (PYR/PYL), which induce phosphorylation activity of the ABA-dependent Sucrose Non-Fermenting Related Protein Kinases (SnRKs) family, and the activation of ABA-dependent transcriptional network involved in ionic and osmotic adjustments in response to salt stress [25,26,27][25][26][27].
Tomato (Solanum Lycopersicum), a widely used vegetable crop throughout the world, is a rich source of antioxidant molecules such as carotenoids, vitamins E and C, ascorbic acid and phenolic compounds, mainly flavonoids. However, salt stress imposes several negative effects on the germination, growth, biomass accumulation and yield of tomatoes [11]. Keeping in view the increasing salinity concerns and global importance of tomato crop, this study was designed with an objective to demonstrate the role of the tomato antioxidative system, RBOH genes, ABA biosynthesis and signaling transcription factor under 12 days long salt stress to provide the basis for future studies regarding the improved salinity tolerance of tomato crops.
2. Salt Stress Enhanced the Activation of Polyamines Metabolism Enzymes in Tomato Seedlings
Under salt stress, the activities of three major polyamine metabolism-related enzymes, ADC, DAO, and PAO, were measured in tomato leaves. The findings revealed that increasing the duration of salt stress significantly increased PA metabolism enzymes’ activity. The activities of two biosynthesizing enzymes, ADC and DAO, and catabolizing enzymes (PAO), increased by 243.42%, 205.00%, and 222.00%, respectively, when compared to the control (Figure 81A–C).
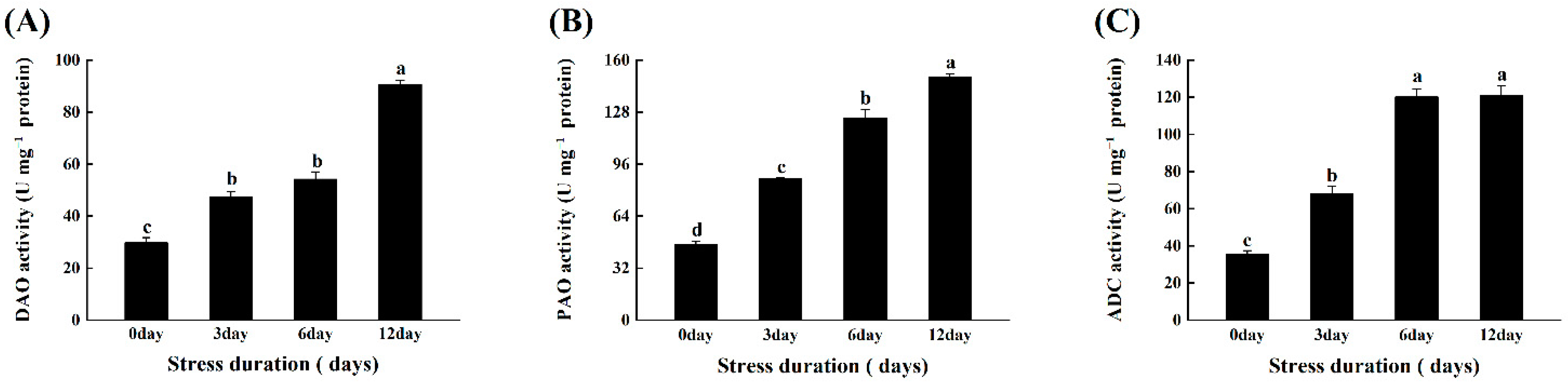
Figure 81. Effects of salt stress on (A) Diamine oxidase (ADC); (B) polyamine oxidase (PAO); and (C) Arginine decarboxylase (ADC) in tomato leaves grown for 12 days under control and salt stress. Values are mean ± SD (Standard Deviation) of three replications. Different letters indicate significant differences at p ≤ 0.05 (Tukey’s HSD test).
3. Salt Stress Upregulated Expression of Antioxidant, RBOHs and ABA Biosynthesis and Signalling Related Genes in Tomato Seedlings
The transcripts abundance of various RBOHs related transcription factors was analyzed through qRT-PCR analysis. The results showed that 12 days of salt stress caused significant up-regulation of SlRBOH1, SlRBOH-A, SlRBOH-D, SlRBOH-E, and SlRBOH-F in the leaves, and SlRBOH1, SlRBOH-A and SlRBOH-D in the roots of tomato seedlings, as compared to the respective control plants. Interestingly the transcription level of RBOH-H in leaves, RBOH-E, RBOH-F and RBOH-H in roots did not show significant variations in their expression levels during the early days (three days in leaves, six days in roots) of salt stress (Figure 92A,B).
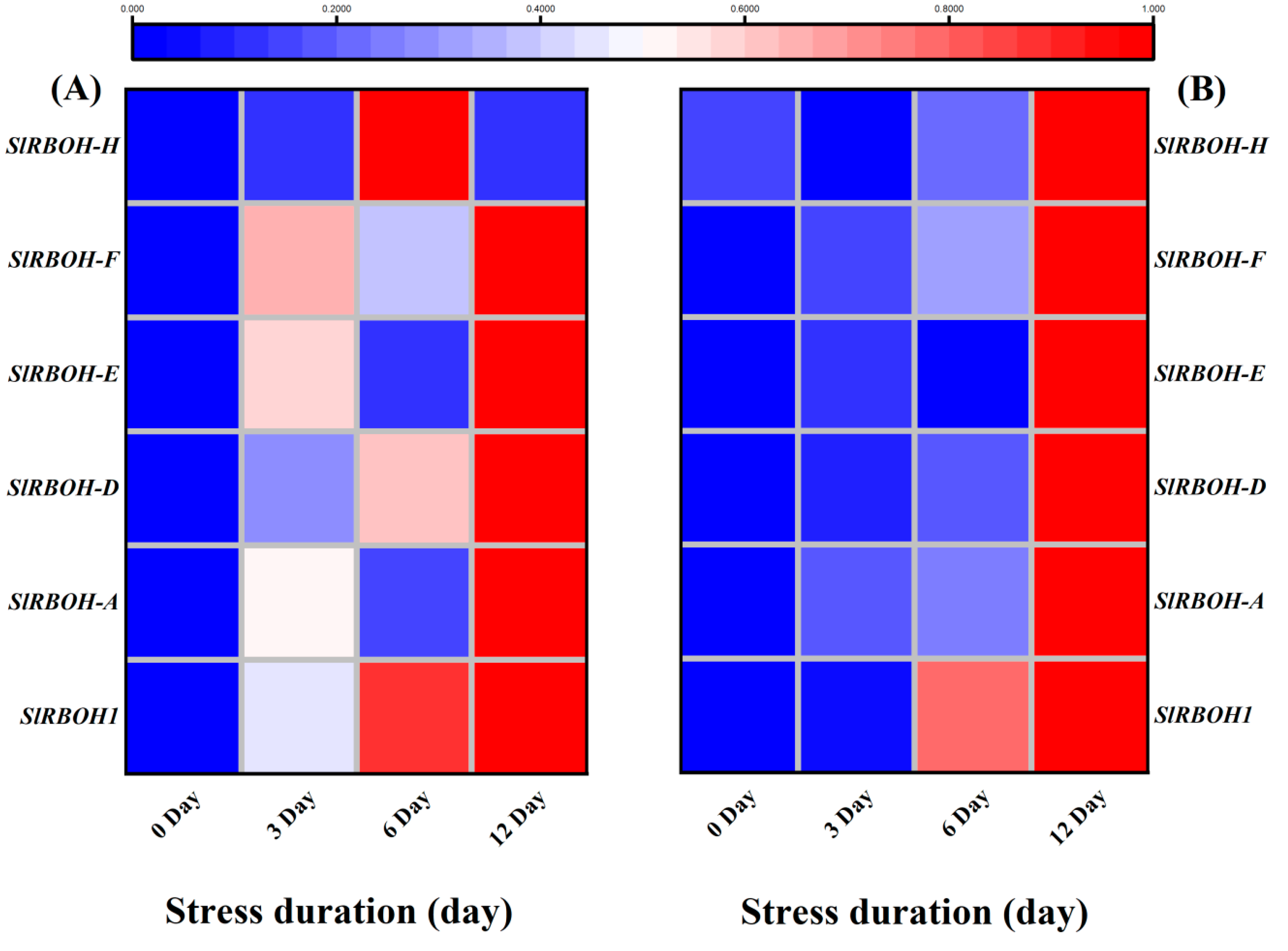
Figure 92. Heatmap of selected genes-related RBOHs (respiratory burst oxidase homolog proteins). The relative expression level was quantified from tomato (A) Leaves; (B) roots after 12days of salt treatment. The scale (Log2 of the mean values after normalization; n = 3) shows the increase in relative concentrations from blue to red color, compared to control (0 h).
In tomato seedlings under salt stress, the ABA content increased with the increasing duration of the salt stress and the highest ABA contents in both leaves and roots was noticed after 12 days of salt stress treatment (Figure 103A,B).
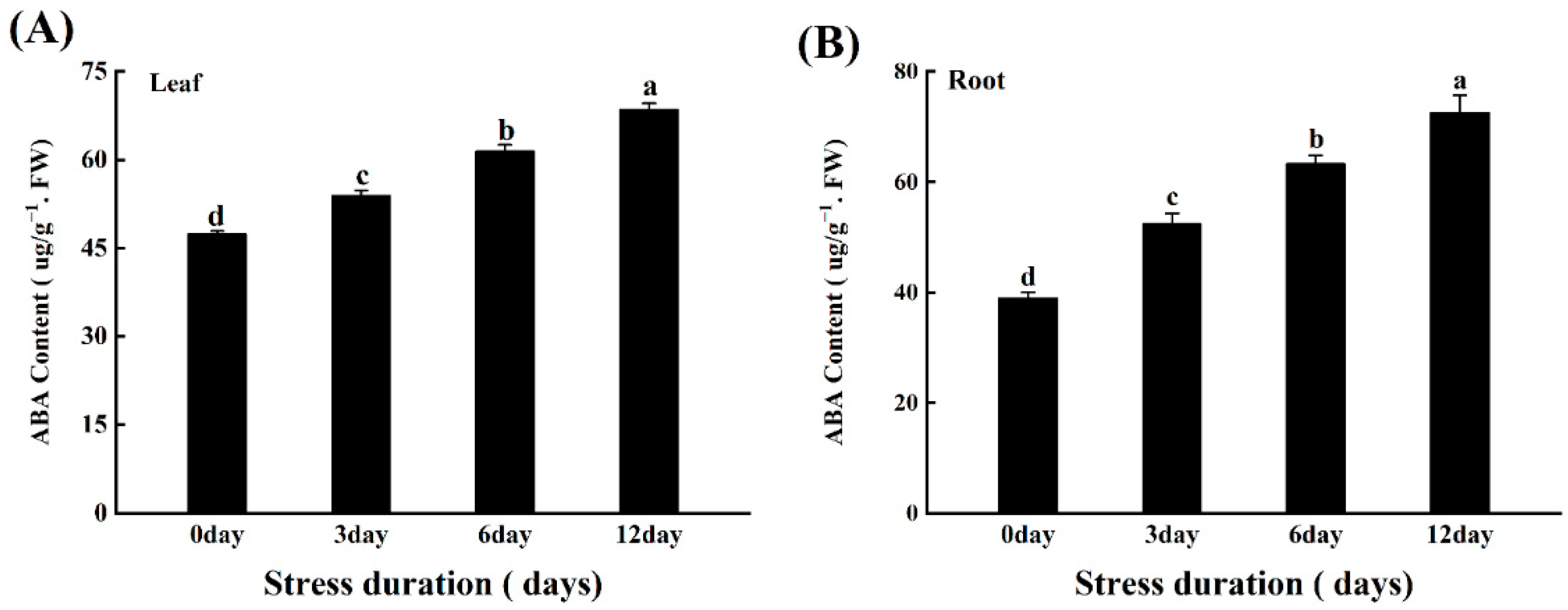
Figure 103. Effects of salt stress on Abscisic Acid in leaves (A) and roots (B) of tomato seedlings. Data was recorded after 12 days of growing tomato seedlings under control and salt stress for 12 days. Values are mean ± SD (Standard Deviation) of three replications. Different letters indicate significant differences at p ≤ 0.05 (Tukey’s HSD test).
Therefore, wthe researchers further analyzed the expression of ABA biosynthesis and signaling genes. Under salt stress, the transcription levels of ABA biosynthesis and signalling genes showed higher expression levels than control. For instance, salt stress upregulated the expression of ABA biosynthesis genes including SlZEP, SlNCED1, SlNCED3, SlNCED5, SlAAO3, SlABI3 and SlABI5 in both leaves and roots, as compared to the control (Figure 114A,B).
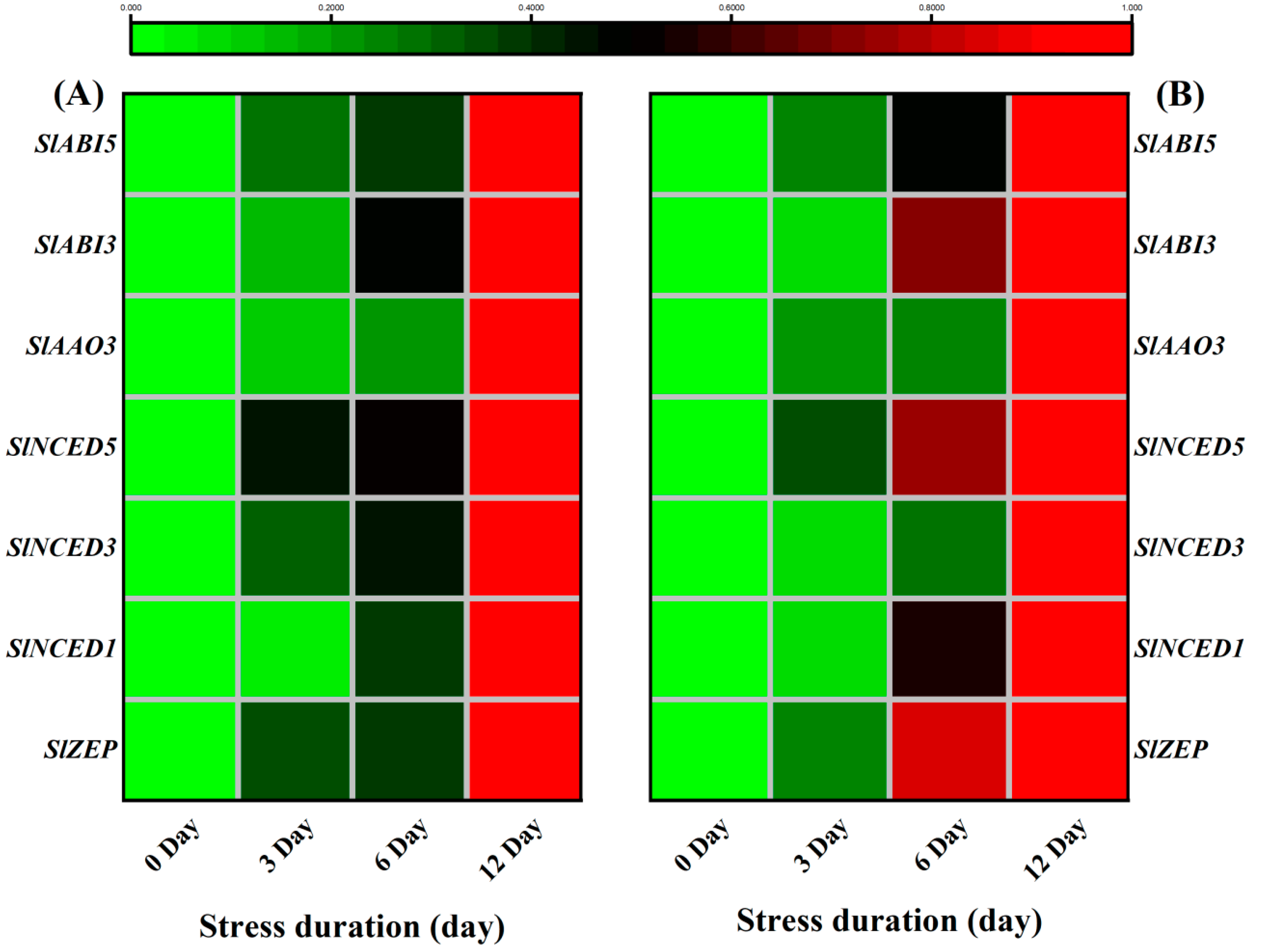
Figure 114. Heatmap of selected genes related to ABA biosynthesis pathway quantified from tomato (A) Leaves; (B) roots after 12 days of salt treatment. The scale (Log2 of the mean values after normalization; n = 3) shows the increase in relative concentrations from green to red color, compared to control (0 h).
Similarly, the ABA signaling genes, SlSnRK2.2, SlSnRK2.3, SlSnRK2.6, SlPYL4, SlPYL8, SlABF4 and SlDREB2 also showed significantly higher transcript levels in leaves and roots in response to salt stress. Taken together, it suggested that the plants tried to counter the damage via enhanced signalling and biosynthesis of ABA (Figure 125A,B).
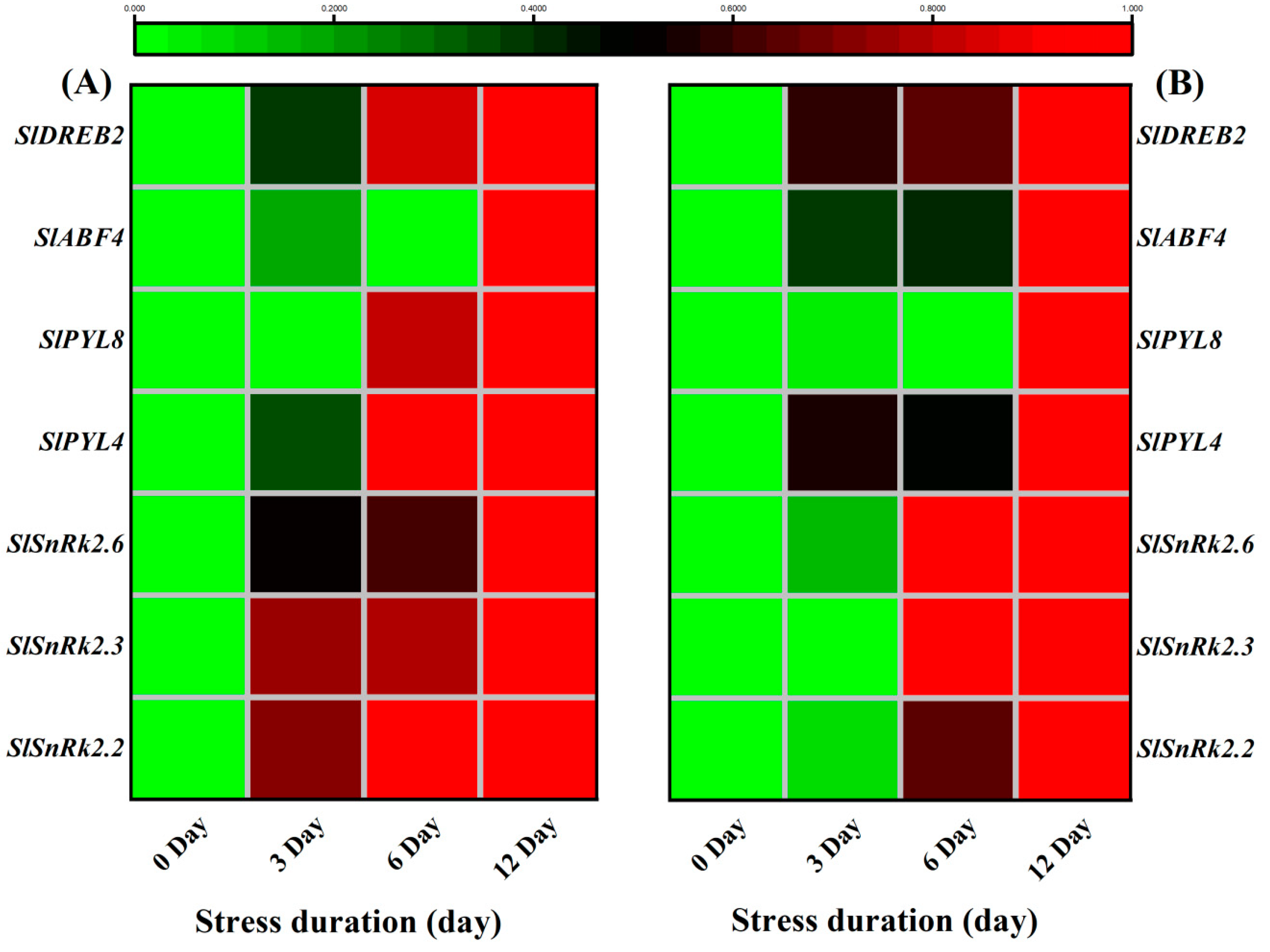
Figure 125. Heatmap of selected genes related to ABA signalling pathway. The relative expression level was quantified from tomato (A) Leaves and (B) roots after 12 days of salt treatment. The scale (Log2 of the mean values after normalization; n = 3) shows the increase in relative concentrations from green to red color, compared to control (0 h).
4. Researches and Findings
Soil salinity is an increasingly severe global problem, as salt hampers plant growth and development and reduces crop yield. In addition to naturally occurring soil salinity, salinization increases due to irrigation practices and climate change [27]. Therefore, in the present study, weresearchers explored the response of the tomato seedlings to three different durations of salt stress. Despite the salt-induced oxidative damage, we the researchers found that tomato seedlings adopted several strategies, including antioxidative scavenging of ROS and activation of stress-responsive gene signalling to detoxify oxidative stress.
One of the most common symptoms of plants under a stressful environment, e.g., salt or drought stress, includes leaf yellowing and growth inhibition, as plants serve their energy on survival rather than growth improvement [28]. Similarly, in the present study, increasing salt stress duration induced significant loss of photosynthetic pigments and markedly affected the morphological indices of the tomato seedlings. Such losses in pigment appear to be caused by the reduced activity of chlorophyll biosynthesis enzymes and increased chlorophyllase activity that degraded the chlorophyll content under salt stress [29,30][29][30]. In addition, sodium ions could disrupt the uptake of some important cation, i.e., Mg+2 that is the main part of the chlorophyll molecule, thus impacting the chlorophyll content of the leaves. Consequently, it impaired the growth and biomass accumulation in the roots and leaves of tomato seedlings. Consistent with ourthe results, number of previous studies found the loss of pigment and growth inhibition of plants under salt stress [7,31,32][7][31][32].
The excess accumulation of sodium ions under the salt stress imposes osmotic stress, disrupting the water permeability and causing a disturbance in the water potential and turgidity of the cells [33]. Consistently, in the present study we also noticed that the values of Ψw and Ψs become more negative, enhancing the corresponding Ψp and osmotic adjustments. Ultimately, the RWC declined significantly. Such reduction in RWC of the cells are the primary indicator of the water stress that limits the water flow to the new cell elongation sites [13]. Similar results were reported earlier in peach [34] and tomato [35].
Moreover, salt stress-induced reduction in the water status of the cell results in stomatal closure, inhibiting the gaseous exchange and photosynthetic ability of the leaves [36]. Combined with the decreased pigment content, it ultimately impairs the photochemical efficiency of the plants. WThe researchers consistently observed decreased values of the gas exchange parameter and maximal photochemical efficiency (Fv/Fm) in tomato seedlings, indicating the severe consequences of salt-induced osmotic stress on tomato seedling photosynthetic efficiency [36,37][36][37]. However, the marked increase in osmotic adjustments in tomato seedlings under salt stress also elucidates that it strives to counter the salinity-imposed osmotic stress by modifying the plant-water relations.
Osmotic regulation is a crucial self-defence mechanism produced by plants under stress. Plants produce high levels of osmoprotectants (e.g., soluble sugars, soluble proteins and proline content) to maintain the osmotic pressure and reduce cell water loss [38]. Proline and soluble sugars accumulated in response to stressful conditions act as a low molecular weight antioxidant that detoxifies the stress-induced toxicity and contributes to cellular osmotic adjustments [39]. In previous studies with various abiotic stresses, including salinity, these osmolytes increased significantly in chickpea [40], Amaranthus tricolor varieties [41], grapes [13] and tomatoes [42]. In addition, burst production of ROS under salt stress could destabilize the protein metabolism, decreasing the contents of soluble proteins in the plants [43]. Consistent with these reports, in our study, protein content decreased. At the same time, proline and soluble sugars were increased with increasing salt stress duration, suggesting that tomato plants suffered salt-induced toxicity and regulated their osmolytes to survive the water limitations under salt stress.
The declined growth parameters and disturbed plant-water relations due to salinity stress indicate a need to study the plant cells’ sodium accumulation and minerals contents.
In the present study, the salt stress obviously enhanced the Na+ accumulation in the roots and leaves of tomato seedlings, disturbing the Na+/K+ ratio and ion homeostasis [44,45][44][45]. It could be due to the activation of the guard cells’ outward rectifying K-channels (GORK), which enhanced the Na+ ion accumulation by the outflow of K+ and resulted in declined K+ levels [44]. Besides the osmotic stress, the higher sodium accumulation in cells could also alter the uptake of other minerals, causing an imbalance in the contents of macro and micronutrients of the plants [45,46,47,48][45][46][47][48]. Similarly, in the present study we researchers noticed a significant decline in the contents of important minerals like P, Mg, Mn and K, which is consistent with the decreased growth and chlorophyll contents of the tomato seedlings under salt stress.
In addition, the significant increase in the contents of heavy metals like Cu and Zn under the salt stress could be attributed to the loss of ion specificity due to excessive salt accumulation in the roots and leaves of tomato seedlings [49]. Previous studies have reported the increase in Ca2+ ions as a response to salt stress to regulate plant response against salt stress [50]. In addition, the wheat genotypes with high salinity tolerance maintained the higher calcium content against salt stress, suggesting the important role of Ca ions in maintaining the ion homeostasis and enzyme activities under salt stress [49]. Consistently, our results also showed higher Ca content in tomato seedlings which could be a response to enduring the salt stress. Collectively, these results showed that salt stress negatively affected the macro and micronutrients in the tomato seedlings. Similar trends were noticed in previous studies with different plant species [51,52,53,54][51][52][53][54].
ROS production, MDA accumulation and electrolyte leakage are the basic biomarkers for oxidative stress under abiotic stresses [55,56][55][56]. In the present study, salt stress induced the production of excessive O2•− and H2O2. Consequently, the higher oxidative stress caused lipid peroxidation and MDA accumulation leading to cell membrane damage and increased leakage of electrolytes as compared to control plants. These results are similar to those in tomato [36], grape [13], and chickpea [40] and demonstrate the severe consequences of salt stress on the growth and development of plants.
Plants have developed different tools against stress which could regulate redox homeostasis and protect plant cells from oxidative damage by scavenging excessive ROS [57]. For instance, SOD is one of the most important antioxidant enzymes that carry out the dismutation of superoxide anion (O2•−) into oxygen (O2) and comparatively less toxic ROS, i.e., hydrogen peroxide (H2O2). Subsequently, the other major antioxidant enzymes like POD, APX and CAT detoxify the H2O2 to water [58,59,60][58][59][60]. In the present study, salt stress-induced activities of various enzymatic (SOD, POD, CAT, APX, GR) and non-enzymatic antioxidants (AsA, GSH) in the tomato seedlings. These results are in line with the previous reports in tomato [30] and other species like Solanum Lycopersicum [36], O. sativa [61], C. arietinum [30], B. juncea [62,63][62][63] and wheat [64]. It suggested that tomato seedlings retained the capacity to tolerate or detoxify the salt-induced toxicity for their survival. Another important antioxidants-mediated protective mechanism in plants includes AsA-GSH cycle that assists the antioxidant enzymes to scavenge the excessive ROS. Both AsA and GSH are potent antioxidants and are considered the buffering agents in redox reactions to protect the plasma membrane from stress-induced oxidation [65]. In this perspective, the H2O2 produced after superoxide dismutation is detoxified by APX into H2O using AsA as the substrate. In the present study, the increased GR activity after salt stress provides GSH, reducing DHAR to dehydroascorbate (DHA) and then to AsA via the AsA–GSH cycle. Similarly, GSH is oxidized to GSSG and subsequently recycled by GR. Thus, the ratio of GSH/GSSG is essential for sustaining the cell redox state [66]. The increase in the activities of SOD, POD, CAT, APX, GR, MDHAR, DHAR, and AsA, DHA was observed in the current study. The ratio of AsA/DHA, GSH, GSSG, and the ratio of GSH/GSSG and proline content in tomato seedlings was observed to be affected by salinity, implying that tomato seedlings attempted to maintain redox regulation under salt stress. It was also confirmed at the transcriptomic level, as the relative transcript levels of antioxidant enzymes showed a similar trend with increasing salt stress duration. A similar gene expression pattern was previously observed in tomato under high temperature stress [67,68][67][68].
In addition, salinity stress also caused a significant increase in polyamines metabolism as suggested by the increase in the activities of three key enzymes named DAO, PAO and ADC. Similarly, in previous reports, salinity stress enhanced the ADC and ODC activities in tomato roots, indicating that both enzymes are responsible for stress tolerance [69]. Furthermore, salinity stress also induced a slight increase in PAO activity [70]. It has been well known that polyamines play important role under stress conditions by modulating ROS homeostasis and regulating the antioxidative mechanism to suppress the ROS production under stress conditions [71]. Therefore, in the present study, the significant increase in enzymes involved in polyamines synthesis could be linked with the signaling regulation to enhance the responsiveness of tomato seedlings against salt stress.
The plasma membrane-localized respiratory burst oxidase homolog (RBOH) proteins generate reactive oxygen species (ROS). AtrbohD and AtrbohF, the major NADPH oxidases responsible for ABA-induced ROS production in Arabidopsis, are involved in stomatal closure [72]. OST1 interacts with AtrbohD and AtrbohF and phosphorylates Ser174 in AtrbohF [73]. To acquire further molecular insights into the salt tolerance mechanism of WT Ailsa Craig, wthe researchers tested the expression of several TFs, ABA biosynthesis enzymes and signalling and defence-related proteins in WT seedlings. Combined with the accumulation of hydrogen superoxide radicals, it could be suggested that salt-induced over-production of H2O2 activated the RBOH signalling mechanism, which regulated the various adaptive mechanisms to counter the salt-induced osmotic stress in tomato seedlings. In previous studies with different abiotic stresses, including low temperature [74], cold [74] and salinity [75], the acclimation to the stresses was found associated with the increased transcript levels of RBOH1 [76]. Furthermore, in Arabidopsis, the expressions of AtrbohD and AtrbohF were upregulated and Na+/K+ homeostasis in wild-type Arabidopsis seedlings that grow on MS medium containing NaCl [77]. Previous studies with sugar beet revealed that BvRBOHE, BvRBOHF, and BvRBOHH downregulates under salt stress. However, wthe researchers found significant upregulation of both SIRBOH-E, SIRBOH-F after 12 days of salt stress and only SIRBOH-H was found downregulated in roots after 12 days of salt stress. It suggests the significant role of RBOH transcription factors in tomato salt-stress acclimatization. Consistent with ourthe results, a genome-wide identification of RBOH genes showed significant upregulation RBOHA, RBOHD, RBOHF and RBOHG under drought stress. Altogether, the significant upregulation of RBOH genes, particularly after 12 days of salt stress, strongly suggests their role in regulating the response of tomato seedlings to long-term salinity stress.
The phytohormone ABA plays a crucial role in regulating a range of plant physiological processes in response to various stresses [78]. ABA functions as an essential secondary signalling molecule to activate a kinase cascade and mediate gene expression during salt stress [79]. Osmotic stress, such as drought and high salinity, dramatically increases the ABA level, which induces the expression of many genes involved in stress responses [80].
In addition to ABA-biosynthesis induction, salt stress also activated the downstream ABA signalling mechanism that controls ABA-regulated gene expression to enhance stress tolerance [86][81]. Various gene families like SnRks [87][82], DREBs [88[83][84],89], PYLs [90][85] and ABFs [91][86] are involved in ABA signalling that improves the tolerance of plants under stressful environments [92][87]. Among SnRKs, Subfamily 2 of SNF1-related protein kinase (SnRK2) is considered as a positive global regulator of abscisic acid signalling [87][82]. Previously, it was noticed that SnRK2.2, SnRK2.6 and SnRK2.3 activated the AREB1/ABF2, AREB2/ABF4, and ABF3 as a response to osmotic stress at the vegetative stage [93][88]. Therefore, the significant upregulation of the ABF4, PYL4 and PYL8 in the present study could also be linked to the higher expression of SnrK genes to improve the ABA-regulated signalling against the salt stress. Consistently, a recent study showed the alterations in transcriptional levels of several salt stress-responsive genes like SlPP2C37, SlPYL4, SlPYL8, SlNAC022, SlNAC042, and SlSnRK2 family by PpSnRK1α, signifying that SnRK1α is involved in the ABA signalling pathway to improve tomato salt tolerance [94][89]. Functional analysis of the AREB/ABFs revealed that these proteins were positive regulators of the ABA-dependent signalling pathways under drought conditions [95][90]. Similarly, physiological, biochemical and transcriptomic analyses showed that SlDREB2 enhanced plant tolerance to salinity by improvement of K+/Na+ ratio and proline and polyamines biosynthesis [96][91].