1. ER+/PR+ Breast Cancers
Estrogen and progesterone hormones influence normal breast development and in breast cancer they play important roles in the disease progression, which is governed by the presence of estrogen and progesterone receptors. The luminal subtypes consist predominately of ER+/PR+ breast cancers and comprise the majority of breast cancers
[1][25]. ER-associated miRNAs are enriched in the luminal subtypes. A key distinguisher between the luminal A and B subtypes is the expression of the Ki67 proliferation marker; luminal A breast cancers have low Ki67, while luminal B breast cancers have high Ki67
[2][46]. The subtype-specific expression of Ki67 reflects the better prognosis and low tumor grade associated with the luminal A subtype, and the comparably worse prognosis and higher tumor grade associated with the luminal B subtype. Furthermore, in terms of miRNA dysregulations,
hwe
re see distinguishing features in the two luminal subtypes. Clustering analysis including breast cancers of all subtypes showed that luminal A and luminal B are clustered close to each other but were distinct from one another
[3][22]. Further, there is more prominent dysregulation in miRNAs in luminal B vs. luminal A breast cancers. Specifically, 657 miRNAs were found dysregulated in luminal B cancers and 67 miRNAs were dysregulated in luminal A cancers. Among the luminal-dysregulation miRNAs, miR-1290 is significantly reduced in luminal A Ki67 low tumors
[4][35]. MiR-1290 is oncogenic in a context driven nature
[5][47]; its reduction in luminal A correlates with its clinical outcome.
The distinct luminal A-miRNA signature is defined by miR-30c-5p, miR-30b-5p, and miR-99a/let-7c/miR-125b cluster, while the luminal B subtype is enriched with miR-182-5p, miR-200b-3p, miR-15b-3p, miR-149-5p, miR-193b-3p and miR-342-3p, 5p
[3][6][7][8][9][22,30,32,33,34]. Haakensen et al. linked miR-30b-5p, miR-30c-5p, miR182-5p and miR-200b-3p to better prognosis in patients with luminal subtype breast cancer
[3][22]. Given the prominence of miR-30 miRNAs among luminal A breast cancers, it important to note that miR-30 is a cluster of six miRNAs (miR-30a, -30b, 30-c1, -30c-2, -30d, 30e), and as whole the miR-30 family generally inhibit migration and growth
[10][48] (
Figure 1). For example, miR-29b/miR-30d regulate migration through lysyl oxidase-like 4 (LOXL4) inhibition
[11][49]. Early evidence demonstrated that miR-30 inhibit cell division through cyclin D2 targeting
[12][50], and inhibition of c-Myc-induced carcinogenesis
[13][51]. The other prominent luminal A-associated miRNA cluster, miR-99a (miR-100)/let-7c/miR-125b), is also typically paired with tumor suppression and reducing growth rate and migration in breast cancer (
Figure 1). This cluster reduces tumor growth by inhibiting proteins involved in important cellular processes, including homeobox A1 (HOXA1), mammalian target of rapamycin (mTOR), insulin-like growth factor binding protein 1 (IGFBP1), and fibroblast growth factor receptor 3 (FGFR3)
[14][15][16][17][52,53,54,55]. The miR-99a cluster also inhibits oncogenes, including Yamaguchi sarcoma viral oncogene homolog 1 (YES1), ETS proto-oncogene 1 (ETS1) and ETS variant transcription factor 6 (ETV6)
[18][23].
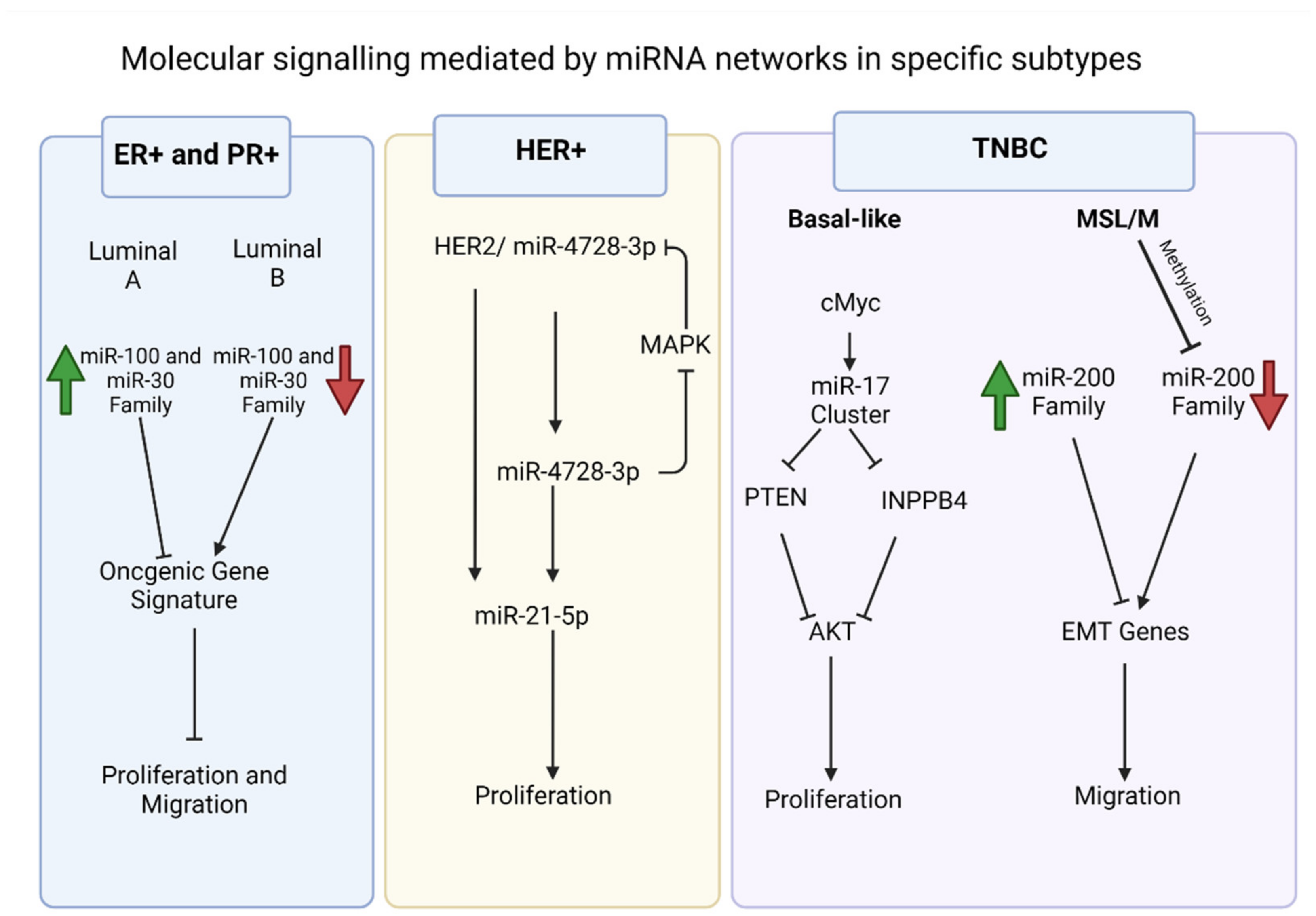
Figure 1. Examples of significant microRNAs (miRNAs) associated with specific breast cancer subtypes and their effects on cell phenotypes. In estrogen receptor positive/progesterone receptor positive (ER+/PR+) breast cancers (blue box), andwe note the role of miR-100 and miR-30 families and the distinction between the luminal A and luminal B molecular subtypes. In human epidermal growth factor receptor 2 positive (HER2+) breast cancers (yellow box), andwe have noted miR-4728-3p, which is present in the intronic region of HER2 and is co-expressed with HER2. The miRNA is involved in feedback regulation of HER2 and oncogenic miR-21-5p. This facilitates several oncogenic processes in later stage tumors. In triple negative breast cancers (TNBCs) (purple box), andwe have noted the cMYC oncogene driven miR-17~92 cluster, which is overexpressed in TNBCs, specifically the BL1 molecular subtype. The miRNA cluster promotes proliferation through its direct targets, which include phosphate and tensin homolog (PTEN) and inositol polyphosphate-4-phosphatase type II B (INPP4B) and are inhibitors of the proliferation mediator AKT. The migratory phenotype of TNBC cell lines that fall within the mesenchymal stem like (MSL) and mesenchymal (M) molecular subtypes is facilitated by inhibition of the miR-200 family through epigenetic changes that allows expression of epithelial to mesenchymal transition (EMT) and migration genes, resulting in the migratory phenotype.
The increased expression of miRNAs associated with tumor suppression among luminal A breast cancers is consistent with the relatively slow growth of luminal A breast cancers; however, there is also evidence that in advanced cancer stages, miR-30 plays an oncogenic role. In patients with advanced tumors, miR-30b-5p is highly expressed in tissue and circulation
[19][38], suggesting a potential change in function towards tumor progression. This is an important caveat when considering miRNA function in cancers; it is always context dependent and reliant on the abundance of relative target mRNAs, which is also shifting during cancer progression. The function of miRNAs is never in isolation and dependent upon the evolving transcriptome.
Additionally, differences in circulating levels of miR-29a, miR-181a, and miR-652 is also evident in the serum samples of luminal breast cancers. All three miRNAs are significantly downregulated in both the tumors and serum of patients with luminal A vs. luminal B breast cancers
[8][33]. Treatment regimens also influence circulating miRNA levels, with tamoxifen treatment of ER+ tumors resulting in increased miR-221 in serum
[20][31].
2. HER2 Overexpressing Breast Cancers
HER2 is a tyrosine kinase receptor that belongs to the family of epidermal growth factor receptors (EGFR). HER2 overexpression in breast cancers is present about 15–20% of breast tumors, and like TNBC, it is associated with worse patient prognosis and survival
[21][56]. However, unlike TNBCs, HER2+ (i.e., HER2 overexpression) in breast cancers can be targeted. Paired with chemotherapy, the monoclonal antibody against HER2, trastuzumab, is used to treat HER2+ breast cancers. However, resistance and recurrence are common, so other drugs have been developed such as the tyrosine kinase inhibitor lapatinib and the monoclonal antibody pertuzumab, which prevents HER2 dimerization and signaling
[22][57]. HER2 overexpression results in specific gene signatures in breast cancer, especially in the cancer-promoting pathways. The increased mitogen-activated protein kinase (MAPK), phosphatidyl-inositol-3 kinase (PI3K)/AKT and HER3 receptor signaling
[23][24][58,59], characteristic of HER2+ breast cancers, result in the downstream effects on enhanced cell proliferation and the observed aggressive clinical phenotype.
HER2+ breast cancers also have a distinct miRNA expression profile. Lowery et al. profiled 453 miRNAs in 29 tumors and found that that the HER2+ subtype is associated with a number of miRNAs, including miR-302c, miR-520d, miR-181c, miR-376b and miR-30e
[25][37]. Additionally, in a study that evaluated 221 breast cancer tumors and 49 normal tissue controls, miR-125b is reportedly upregulated in HER2+ breast cancers
[26][36]. MiR-4728-3p expression is also associated with HER2+ breast cancers and its gene is encoded within a HER2 intron
[27][60]. Other miRNAs identified in HER2+ breast cancers include upregulation of miR-21 and miR-146a-5p, while miR-181d and miR-195-5p are downregulated
[28][61]. These altered miRNA expression profiles are specific to the HER2+ subtype and could be exploited in diagnostic/prognostic tools.
In terms of a connection with cancer phenotypes associated with the HER2+ breast cancer subtype, these miRNAs also have function beyond just expression associations. For example, miR-125b is specifically connected to metastasis of HER2+ breast cancers
[29][62] and with worse patient outcomes
[30][39]. However, the effects of the miRNA in cancer are again context dependent, as miR-125 has also been reported to have tumor suppressive effects in various cancers
[31][63]. For HER2+ associated miR-4728-3p, its mRNA targets include downstream targets of HER2 signal transduction and the estrogen receptor alpha (ESR1)
[32][64]. Recent investigation into miR-4728 indicate that when it is overexpressed in HER2-postive tumors, the efficacy of HER2 inhibitor laptinib is minimized (
Figure 1)
[33][65]. This was linked to decreased expression of pro-apoptotic NOXA59. Further, miR-4728-3p mediates stabilization of miR-21-5p in HER2+ breast cancer (
Figure 1)
[34][66], facilitating miR-21-5p mediated oncogenesis
[35][36][67,68].
As demonstrated with miR-4728
[33][65], the effect of miRNA on drug treatments are not uncommon, likely because miRNAs are expressed in a context dependent manner. Identifying changes to the miRNA landscape before and after treatment will aid in the development of improved treatments for HER2+ breast cancers. Normann et al. utilized four HER2+ breast cancer cell lines to assess whether treatment with trastuzumab and lapatinib together or separately change the miRNA landscape
[22][57]. Levels of miRNAs hsa-let-7b, miR-1236, miR-134, miR-25, miR-3656, miR-3663-3p, miR-3940 and miR-885-5p were significantly altered following drug treatment
[22][57]. Importantly, treatment with miRNA mimics (e.g., miR-101-5p mimic) sensitized cells to treatment with the drugs. Additionally, miR-101-5p downregulates HER2 and MAPK1; its expression is associated with higher survival rates in HER2+ tumors. These findings together indicate that miR-101-5p acts as a tumor suppressor in HER2+ cancers.
3. TNBC
TNBC is one of the major contributors of breast cancer mortality; nearly 25% of the overall breast cancer-related deaths are among patients with TNBC, despite representing only 10–15% of breast cancers
[37][40]. TNBCs are aggressive in terms of proliferation, with a high mitotic index and high Ki67 staining in histology. They also recur more frequently than other breast cancer subtypes. Like the other above reviewed subtypes, TNBC also has reported correlations with specific miRNAs. For example, survival analysis of patients reveal correlation with multiple miRNAs in TNBC. Specifically, miR-27a/b, miR-210 and miR-30e are associated with worse survival, and miR-155 and miR-493 are associated with better survival in TNBC
[38][39][40][41,42,43]. Further, miR-374a/b and miR-454 are associated with disease free survival
[32][64]. Some miRNAs are reported to be associated with metabolic processes in TNBC; miR-210, miR-105-5p and miR-767-5p are essential for the Warburg effect, with miR-210 involved in glucose uptake, lactate production and extracellular acidification rate in TNBC
[40][43].
TNBCs exhibit significant heterogeneity, both phenotypically and genotypically
[41][42][44,69]. Hence, TNBCs can be further subdivided into multiple subtypes. Based on gene expression profiling, TNBC is divided into six major subtypes; basal-like 1 (BL1), basal-like 2 (BL2) immunomodulatory (IM), claudin-low mesenchymal (M), mesenchymal-stem like (MSL) and luminal androgen receptor positive (LAR), and each of these have unique clinical outcomes, phenotypes and drug sensitivities. For example, the two mesenchymal TNBC subtypes are associated with epithelial–mesenchymal–transition (EMT) gene signatures and pronounced migratory capacity. The individual miRNA signature for molecular subtypes has not been studied in detail for most of the TNBC molecular subtypes. However, significant information on basal-like TNBC are achieved through in silico analysis of patient cohort studies with miRNA studies. The molecular influences of some of the miRNAs are studied in detail in in vitro experiments detailing its function in TNBC.
For example, Rinaldis et al. identified a distinct four miRNA signature in TNBC and demonstrated that the miR-17~92 and miR-106b-25 clusters are significantly overexpressed in basal-like TNBCs
[43][70]. It is noteworthy that the miR-17~92 cluster is of particular importance in the TNBC transcriptome and are among the most well-studied miRNAs (
Figure 1). The miR-17~92 cluster comprises of miR-17, miR-18a, miR-19a, miR-20a, miR-19b-1 and miR-92a-1; together these miRNAs regulate expression of transcription factor E2F1, thrombospondin1 (THBS1), connective tissue growth factor (CTGF) and phosphate and tensin homolog (PTEN)
[44][45][46][71,72,73]. Earlier, O’Donnell et al. showed that the proto-oncogene cMYC modulates the critical transcription factor E2F1 by regulating the miR-17~92 cluster, resulting in cancer proliferation
[47][74]. This intricate oncogene signaling regulation has earned the miR-17~92 cluster the moniker oncomiR-1. Apart from involvement in cMYC oncogene signaling, the oncomiR-1 cluster also inhibits inositol polyphosphate-4-phosphatase type II B (INPP4B) and is distinctly associated with the BL1TNBC subtype
[48][75]. Notably, INPP4B is a known inhibitor of the PI3K/AKT mediated growth pathway; knockdown experiments clearly showed the tumor suppressive role of INPP4B
[49][76]. Further, Kalecky et al. found distinct differences between BL1 and BL2; miR17~92 cluster, miR-17, miR-18a and miR-19a were high in BL1 tumors, but not as high in BL2 tumors
[48][75].
The BL1 subtype of TNBC is associated with low PTEN and overall decreased miRNA expression
[50][77]. This group is also associated with worst survival among TNBCs. Recent evidence on structural mapping of miR-17~92 cluster showed a sub-optimal micro processing of primary miRNAs, resulting in unequal expression of the constituent miRNAs of the cluster
[51][78]. This highlights the potential role of regulatory molecules of miRNA processing in the regulation of the miR-17~92 cluster
[52][53][79,80]. This cluster may play a crucial role in the subtype characteristics associated with BL1-TNBCs and the distinction of TNBCs from ER+/PR+ and HER2+ breast cancers.
Like the miR-17~92 cluster, miR-135b shows molecular subtype specificity among TNBCs; miR-135b is upregulated in TNBCs in general but is most overexpressed in BL1 and BL2 TNBC subtypes
[54][81]. Importantly, miR-135b regulates expression of the ER, androgen receptor (AR) and hypoxia inducible factor 1 alpha subunit inhibitor (HIF1AN)
[55][82]; thereby, its expression in TNBCs may directly affect hormone receptor loss and contribute to this distinguishing feature of TNBCs. Interestingly, the AR significantly reduces miR-135b expression
[54][81]. Further, miR-135-b also targets large tumor suppressor kinase 2 (LATS2), a intermediator of Hippo signaling, and promotes cell proliferation in unstratified breast cancer patient tumors
[56][83]. In TNBC, miR-135-b targets adenomatosus polyposis coli (APC), a WNT signaling regulator
[57][84]. Both these targets are important regulators of migratory process. These studies observed increased proliferation and migration in the cells treated with miR-135b mimics. However, in early-stage breast cancer, miR-135b-5p repression resulted in migration through the syndecan-binding protein (SDCBP)
[58][85]. These findings strongly suggest that miR-135-5p has a role in the regulation of migration processes in breast cancer, with functional difference among different subtypes arising from context-specific signaling networks.