‘Lamon bean’ is a protected geographical indication (PGI) for a product of four varieties of bean (Phaseolus vulgaris L.) grown in a specific area of production, which is located in the Belluno district, Veneto region (N.E. of Italy). TIn the last decade, the ‘Lamon bean’ has been threatened by severe virus epidemics that have compromised its profitability. TIn this work, the full virome of seven bean samples showing different foliar symptoms was obtained by MinION sequencing. Evidence that emerged from sequencing was validated through RT-PCR and ELISA in a large number of plants, including different ecotypes of Lamon bean and wild herbaceous hosts that may represent a virus reservoir in the field. Results revealed the presence of bean common mosaic virus (BCMV), cucumber mosaic virus (CMV), peanut stunt virus (PSV), and bean yellow mosaic virus (BYMV), which often occurred as mixed infections. Moreover, both CMV and PSV were reported in association with strain-specific satellite RNAs (satRNAs). In conclusion, this work sheds light on the cause of the severe diseases affecting the ‘Lamon bean’ by exploitation of MinION sequencing.
- plant disease
- virology
- bean common mosaic virus
- cucumber mosaic virus
- peanut stunt virus
- bean yellow mosaic virus
1. Introduction
Considering the wide diversity of species and viral strains that could be detected in Lamon bean, a fully comprehensive and sensitive diagnostic tool, able to explore such a great variability, was highly needed.
For this reason, rwesearchers took advantage of the advent of High Throughput Sequencing (HTS) approaches, which in the last two decades have allowed great advances and have pro- moted discovery, diagnostics, and evolutionary studies [3][20]. Plant virus research has been heavily impacted by the development of HTS techniques for the identification of emerging viruses, genome reconstruction, analysis of population structures, evolution of novel viral strain(s), and much more [4][5][6][7][8][9][21–26]. Single-molecule sequencing technologies, often called “third generation sequencing”, provide greater advantages over second-generation ap- proaches, including the production of long read lengths that are easier to map to a reference sequence and that facilitate de novo assembly, short run times, small amounts of input nu- cleic acids (DNA or RNA), and low cost for a single run [3][20]. The consistent improvement in HTS technologies has occurred in parallel with the continuous development of suitable bioinformatic tools for data analysis and with the online availability of a wide range of biological data sets, which have allowed a deeper comprehension of HTS results [6][7][23,24].
In this entworyk, a third-generation sequencing platform, namely MinION, provided by Oxford Nanopore Technologies, was exploited to obtain the virome of seven bean samples that exhibited different foliar symptoms resembling those of virus disease. The picture that emerged from the sequencing results was consolidated through RT-PCR and ELISA assays in many samples, inclusive of different varieties of bean grown in the Lamon area and various families of herbaceous plants, which may represent a virus reservoir in the field.
2. Virome Determination
2.1 Virome Determination

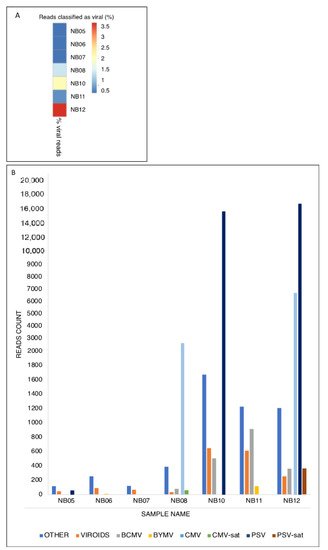
3
2.2. RT-PCR and ELISA Detection
A total of 59 samples were tested by RT-PCR assays to confirm the presence of the viruses detected by MinION sequencing, BCMV, CMV, PSV and BYMV (Table 1). Three Lamon beans were negative to all tested viruses (9%). Single infections caused by BCMV, and PSV were detected in 39% and 13% of Lamon beans, respectively, while CMV was reported in 21% of bean samples, exclusively in co-presence with either BCMV (9%), PSV (3%), or both (9%). Mixed infection by BCMV and PSV was also reported in 12% of Lamon beans. BYMV was identified, exclusively by MinION sequencing, in two samples (6%) either as a single infection (LB-21, NB06) or in co-presence with BCMV (LB-4, NB11). All herbaceous hosts were negative for the presence of BCMV. CMV was exclusively reported in Trifolium pratense L. (Leguminosae), both as a single infection (4%) and in co-presence with PSV (8%). Single infection caused by PSV was detected in Trifolium pratense L. (8%), Chrysanthemum sp. (4%) and, for the first time, in Solanum tuberosum Solanum tuberosum L.L. (8%). Members of the Amaranthaceae (Chenopodium album L., Amaranthus retroflexus L., and Achillea millefolium L.), Apiaceae (Daucus carota L.), and Asteraceae (Taraxacum officinale L.) were negative to all tested viruses (68%). The results of ELISA tests are reported in Table 2 and revealed BCMV presence in samples collected in all the surveyed municipalities, with an incidence ranging from 40% to 100%. CMV was serologically identified in 50% and 62% of the samples belonging to the two municipalities (Sovramonte and Belluno) with incidence of 25 and 30%, respectively.
4. BLAST Analysis of Sanger Sequences and Phylogenetic Investigation
2.3. BLAST Analysis of Sanger Sequences and Phylogenetic Investigation
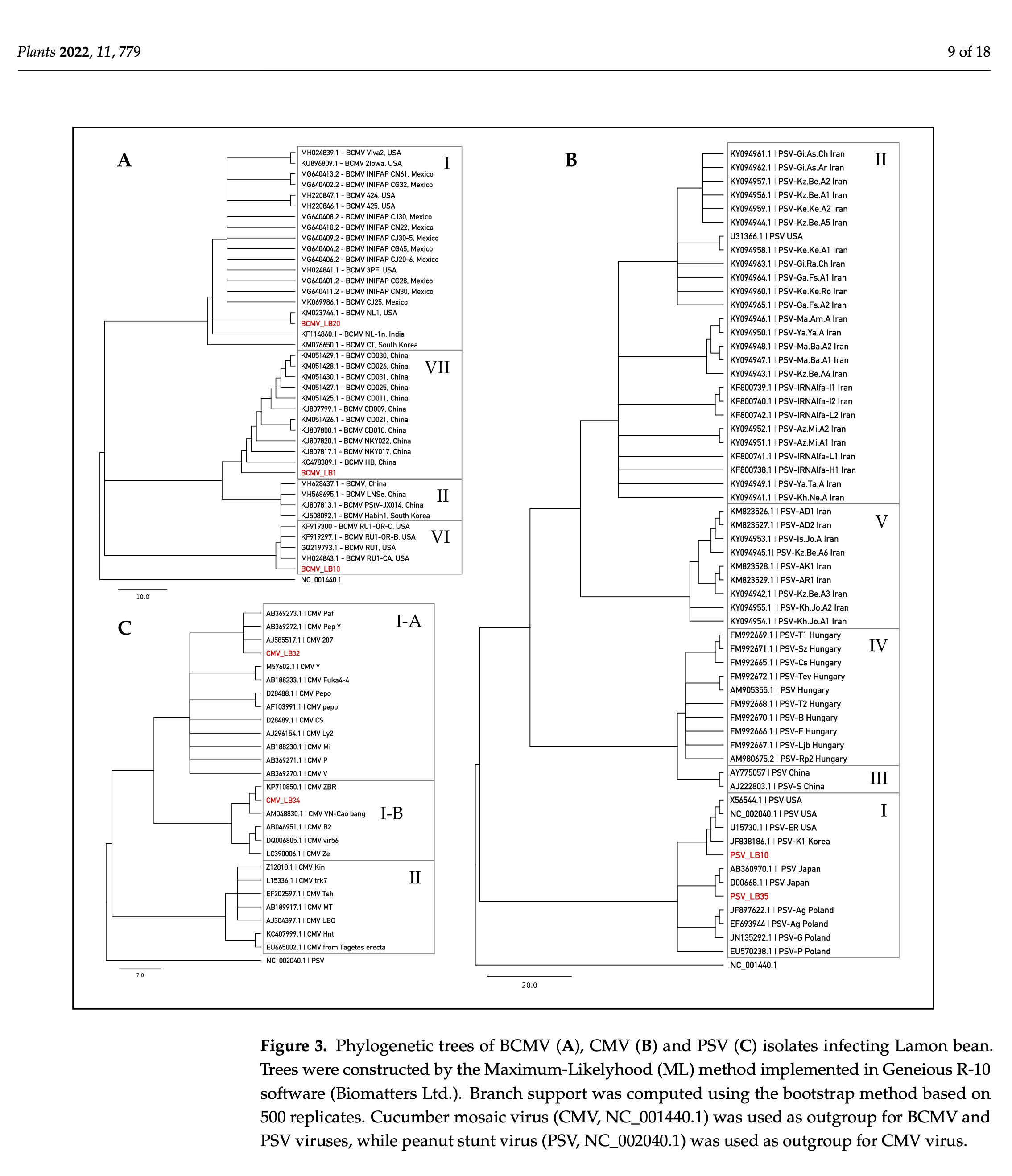
Sequences obtained by Sanger were used for assignment to pathotypes/subgroups according to recent comprehensive studies of BCMV [10][27], CMV [11][28], and PSV [12][29]. The results revealed that 79% of BCMV sequences fall into pathotype VII, sharing 97.7–99.2% identity with NL-4 strain (DQ666332) [13][30]. Three BCMV sequences (13%) were in- cluded in the pathotype I, exhibiting from 99.0% to 99.6% of sequence similarity to the NL-1 strain (KM023744) [14][31]. The remaining BCMV sequences (8%) fall into pathotype VI, sharing 95.6–99.2% identity with recombinant RU-1 (GQ219793) [15][32] and RU-1-CA (MH024843) [16][33] strains. All PSV sequences belonged to subgroup IA, showing a se- quence similarity that ranged from 92.7% to 99.3% with the PSV-ER strain (U15730) [17][34]. CMV sequences can be assigned to the subgroups IA and IB, exhibiting 99.3% iden- tity with Fny-CMV strain (D10538) and 95.3–96.4% sequence similarity with As-CMV strain (AF013291), respectively [18][19][20][10,35,36]. Results of BLAST analyses are summarized in Table 3 Table 3. Phylogenetic analyses (Figure 3) assigned reference sequences of the isolates of BCMV, PSV and CMV detected in Lamon bean plants to the respective clusters congru- ently with the BLAST results outlined above. The phylogenetic analysis carried out with the introduction of other sequence variants found in the survey produced similar results (not shown).

5. Discussion
An all-encompassing description of the virome associated with a crop and the availability of high-throughput diagnostic tools are useful to improve viral disease control strategies. The continuous development of newer and even more robust and user-friendly High Throughput Sequencing (HTS) technologies for detecting and identifying viruses makes these new approaches suitable for routine use in testing laboratories, representing a powerful new high-throughput diagnostic tool [3][21].
Moreover, to assess confidence on MinION sequencing as a diagnostic tool for laboratory routine diagnosis, as well as on its reliability in the identification of novel viral strains, Sanger sequences assigned to BCMV, PSV, and CMV were compared to HTS contigs obtained by de novo assembly of reads assigned to corresponding viruses by our bioinformatic analysis. Alignments revealed that BCMV (Figure S1), PSV (Figure S2), and CMV (Figure S3) contigs obtained assembling MinION reads showed 100%, 99.5%, and 99.1% sequence similarity with the corresponding Sanger sequences, respectively.
3. Discussion
An all-encompassing description of the virome associated with a crop and the avail- ability of high-throughput diagnostic tools are useful to improve viral disease control strategies. The continuous development of newer and even more robust and user-friendly High Throughput Sequencing (HTS) technologies for detecting and identifying viruses makes these new approaches suitable for routine use in testing laboratories, representing a powerful new high-throughput diagnostic tool [20,37].
In this work, MinION sequencing by Oxford Nanopore Technologies was exploited as a diagnostic method for detection and identification of viruses infecting Lamon bean, an agricultural product with protected geographical indication.
MinION sequencing provides many advantages over the widely used RNA-seq ap- proach. Minion platform produces long read lengths, which are easier to map to a reference genome, and facilitate de novo assembly. In addition, this approach significantly reduces run times, requires only a small amount of total RNA, and has low costs for a single run. The MinION approach was preferred to the commonly used RNA-seq, because it can be easily performed in every laboratory using the simple and small sequencer provided with the kit, and data analysis for diagnostic purposes does not necessitate deep knowledge of bioinformatics tools [20].
Overall, a single MinION sequencing run allowed a snapshot of the viromes of seven samples, enabling a high-throughput screening of their infection status. Moreover, since the samples were ligated without the use of PCR, the number of viral reads assigned to each sample can be directly linked to the viral load. Indeed, the percentage of viral reads in our samples suggested a wide range of viral loads. The virome analysis of seven bean samples, which exhibited different foliar symptoms, revealed the main presence of BCMV, CMV, PSV, and BYMV, which occurred both singly and in mixed infection [38–41].
RT-PCR assays detected BCMV in many bean samples (75%), whereas it was never reported in herbaceous hosts, thus excluding their possible roles as virus reservoirs in the field. Sequence analyses and phylogenetic investigations performed on a 505-bp fragment of BCMV polyprotein revealed that the strains found in the Lamon area fall into pathotype I and VII, showing high similarity with the NL1 (KM023744.1) and NL4 (DQ666332.1) strains, respectively [30,31]. Two BCMV sequences have been included in pathotype VI, exhibiting significant sequence similarity to RU-1 (GQ219793) and RU-1-CA (MH024843), which have been reported as capable of overcoming bc−22, the most advanced resistance gene in the common bean [32,33,42].
Our data revealed a wide intra-specific genetic variability in the BCMV population that led to a challenging visual assessment of symptoms in infected plants, thus complicating the identification of putative tolerant genotypes. The presence of BCMV isolates that showed high sequence identity with the RU-1 and RU-1-CA strains suggests that the phenomenon of overcoming resistance may also occur [32,33]. Moreover, the presence of mixed infections with different viral species further interferes with evaluation of the type and severity of disease symptoms.
Molecular diagnosis revealed the presence of the cucumovirus CMV, and its associated satellite RNAs (satRNAs) in 25% of the Lamon beans and 7% of the herbaceous hosts tested. The virus was often detected in co-presence with BCMV. Sequence analysis and phylogenetic studies performed on a 584-bp fragment encoding for the coat protein (CP) gene of CMV demonstrated that viral isolates infecting bean in the Lamon area are genetically similar to the Fny and Cs strains, clustering into subgroups IA and IB, respectively [43]. The CMV-associated satRNAs identified in Lamon bean showed high sequence similarity to the T4, T8A and IR-WI classes, as well as the WL3-satRNAs, pBsat2 RNA, and E-satRNA.
In viral diseases, satRNAs play pivotal roles in symptom expression, by worsening, attenuating, or modifying symptoms in infected plants through different molecular mecha- nisms [44,45]. The common effect of satRNAs is the attenuation of CMV-related symptoms, which occurs often (but not always) through a noticeable decrease in accumulation of viral RNAs [46,47]. This effect is due to the competition between virus and satRNAs for shared replicase-related factors [47,48]. Different classes of CMV-satRNAs can produce distinct phenotypes on infected plants, depending on both the host and the helper virus [49]. Moreover, the presence of certain satRNAs may also affected the pathogenicity of CMV isolates, depending on whether the disease symptoms are unaffected, exacerbated, or attenuated by the presence of satRNA [50].
The classes of satRNAs detected in Lamon beans have been suggested to attenuate CMV-related symptoms [45,51–53], despite there still being a lack of experimental evidence. The synergy between CMV and potyviruses, such as BCMV, has been well-characterized, mostly (but not exclusively) in cucurbit hosts [38]. Wang et al. demonstrated that in CMV-infected plants, symptom attenuation mediated by CMV-satRNAs was suppressed in the presence of potyviruses such as BCMV [38].
Thus, the co-presence of BCMV and CMV, which has been frequently reported in the Lamon area, could further interfere with symptom expression. Indeed, we can speculate that the presence of BCMV in CMV-infected bean could suppress the beneficial effect of CMV-satRNAs, determining an increase in the CMV concentration in infected tissues, and resulting in worse disease symptoms.
PSV was reported in 33% of the Lamon bean samples, representing the first report of the virus in Phaseolus vulgaris L. in Italy. The presence of this virus in Lamon bean was also confirmed by ELISA. PSV was also detected in 22% of samples from herbaceous hosts. The viral strains detected in Lamon can be ascribed to subgroup I, showing high sequence identity with the PSV-J strain and PSV-ER strain. The PSV-associated satRNA showed significant sequence similarity with the P-satRNA strain, which was reported to be involved in worsening of disease symptoms in PSV-infected plants [54]. As demonstrated for CMV satRNAs, PSV satRNAs are also competitors for the same helper-virus replication machinery, thus determining a decrease in viral concentration in infected tissues [55]. Plant proteome changes in response to the presence of P-satRNA during PSV infection have been investigated, demonstrating that P-satRNA induces significant decrease in levels of proteins involved in carbohydrate metabolism, protein biosynthesis, and stress-related factors, including aminopeptidase protein, which is the satRNA-responsive factor [56]. Interestingly, for the first time PSV has been reported in two plants of Solanum tuberosum cultivated nearby a bean patch in the Lamon municipality. PSV isolates collected from S. tuberosum were sequenced by Sanger technology, revealing a significant sequence identity (>97.0%) with PSV-ER type strain, clustering into subgroup IA together with the isolate PSV_LB35 (accession OL875025) recovered from Lamon bean plants (data not shown). Further studies including a great number of samples need to be performed to evaluate genetic variability and viral pathogenesis on this novel host.
Our analysis based on MinION sequencing showed that BYMV was detected in two of our samples, but this result was not confirmed by RT-PCR. Two different primer pairs were employed in conventional PCR: the first primer pair was retrieved from the literature [57], while the second one was newly designed for this study. However, neither of these primer pairs were able to detect BYMV in the Lamon beans analysed in this study. We investigated why this may have occurred and discovered that the sequenced strains shared only approximately 90% sequence identity with the BYMV reference sequence. This observation may suggest the appearance of new viral strain(s) [26,41], excluding high sequencing error rate, often ascribed to the MinION technology.
In the past, the high sequencing error rate has been frequently reported for MinION technology, representing a significant weakness of this strategy [58]. Recently, this drawback has been largely mitigated, thanks to the development of new sequencing chemistries and a new generation of highly accurate base-callers, such as Guppy [59]. Given the high proportion of MinION detected viruses validated with gold-standard methods, this novel protocol represents a reliable approach for the rapid and high-throughput detection of viruses in a large number of samples, including different ecotypes of Lamon bean and various herbaceous hosts. This aspect was further corroborated by comparing Sanger sequences of BCMV, PSV, and CMV with the corresponding sequences obtained through HTS approach.
Overall, our results suggested a wide inter- and intra-specific variability among viruses detected in Lamon beans, revealing the existence of mixed infections that may promote viral interactions, such as synergisms or antagonisms [60,61]. Moreover, the survey is also complicated by the presence of CMV- and PSV-satRNAs, for which confirmation of their involvement in plant responses and symptom expression needs to be done.
The results obtained in this work represent a fundamental approach aimed at de- scribing the viruses involved in the Lamon bean pathosystem. In a context of sustainable management of Lamon bean IGP production, a strategy of selection under epidemic pres- sure of individuals showing tolerance to viruses may be worthy of consideration. Moreover, the comprehensive knowledge of viruses affecting Lamon beans will help to improve the management strategies based on virus-free propagation material and on chemical control of aphid vectors.