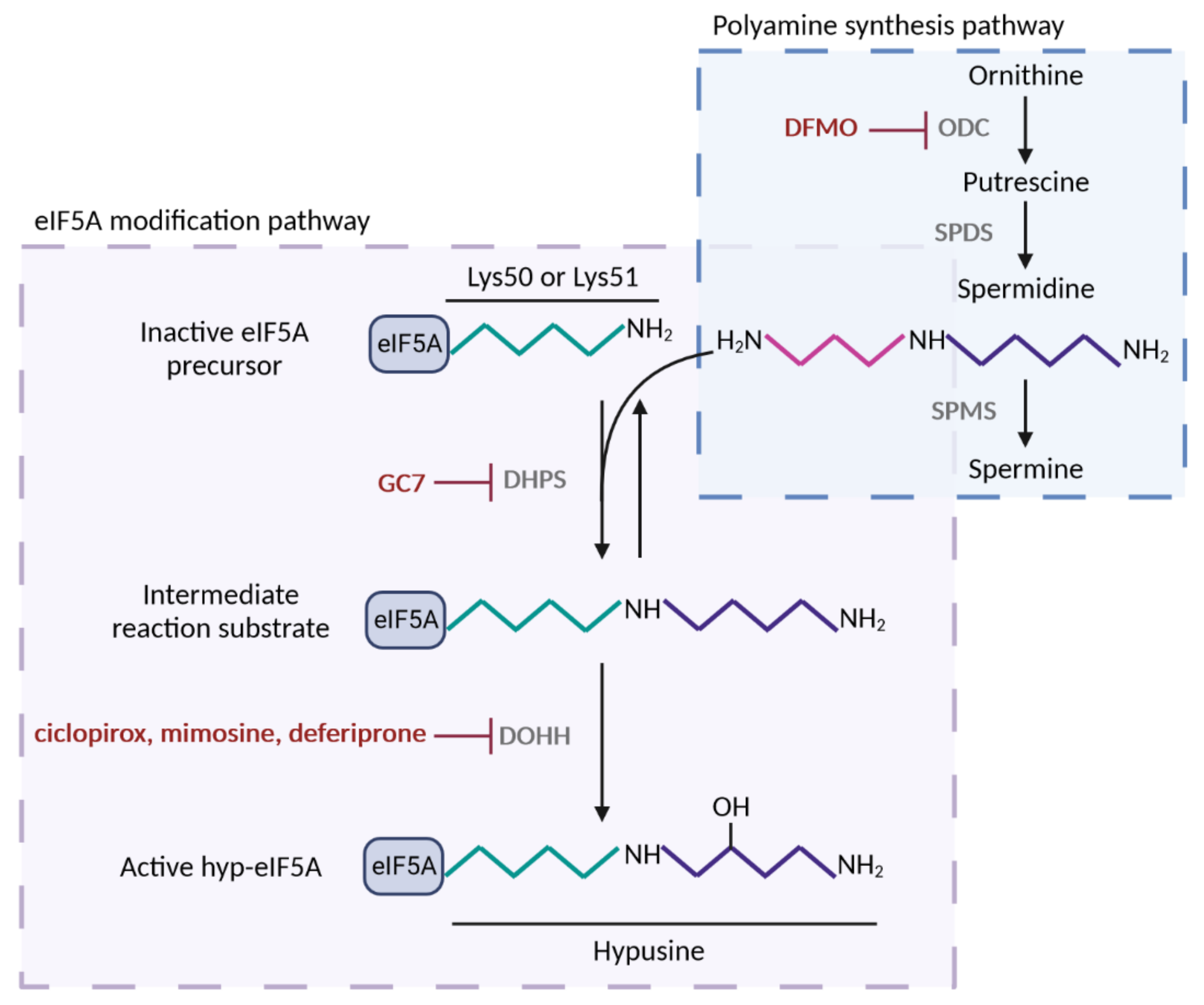
Figure 1. Polyamine-hypusine pathway and its pharmacological inhibitors. Spermidine substrate for eIF5A hypusination is obtained by the conversion of the polyamine ornithine in putrescine by the enzyme ornithine decarboxylase (ODC); next, spermidine is synthesized from putrescine by spermidine synthase (SPDS). Alternatively, spermidine is converted in spermine by spermine synthase (SPMS). Hypusine modification of lysine-50 (human) or lysine-51 (yeast) residue of eIF5A occurs by the addition of spermidine via two consecutive enzymatic reactions. First, deoxyhypusine synthase (DHPS) transfers the aminobutyl group of spermidine to the amino group of lysine generating an intermediate substrate, which does not accumulate. Second, deoxyhypusine hydroxylase (DOHH) adds a hydroxyl group and forms the hypusine residue of eIF5A, which confers the activity to the protein. eIF5A post-translational modification can be suppressed by inhibitors of DHPS and DOHH, but also by inhibition of ODC, the rate limiting enzyme for spermidine biosynthesis. Figure processing was carried out using BioRender software.
Regarding the structure of eIF5A, different studies, whose subjects range from archaea to humans, have been published in the last decades
[16][17][18][16,17,18] showing how hyp-eIF5A folds into a two-domain structure of predominantly β-sheet character, in which the N-terminal portion harbors the unique feature of eIF5A, the hypusine residue
[19]. This residue is located at the tip of an extended, unstructured, and exposed loop (hypusine loop) resembling a tRNA. After binding to the already-formed 80S ribosomal complex, hyp-eIF5A is predicted to lie adjacent to the P-site tRNA overlapping the E-site
[20][21][22][20,21,22]. In this way, hyp-eIF5A prevents ribosomes from stalling at specific sequences by projecting the hypusine-containing domain toward the P-site to sterically restrict the position of the residue placed on it. Specifically, hyp-eIF5A stimulates the synthesis of proteins by favoring peptide bond formation between critical amino acid residues known to be poor substrates for the reaction, such as stretches of three or more consecutive proline residues (PPP) or polyproline motifs, but also combinations of proline, glycine, and charged amino acids
[9][10][11][9,10,11]. Thus, hyp-eIF5A assists in the translation of only a part of the overall mRNA population, which is its distinctive feature. In connection with its main role in translation, hyp-eIF5A can also be localized to the endoplasmic reticulum (ER), where it is associated with ribosomes bound to the ER membrane, and it seems to facilitate the co-translational translocation of some proteins into the ER, such as collagen
[23][24][25][26][23,24,25,26]. Thus, blocking eIF5A hypusination upregulates the stress-induced chaperones in yeast
[25] and leads to ER stress in mammalian cells
[26][27][26,27].
Several studies have indicated that eIF5A is involved in processes that are not directly related to protein synthesis. The structural features of eIF5A suggest it offers the potential to interact with nucleic acids. The C-terminal domain resembles the cold-shock domain (CSD), common in DNA- and RNA-binding proteins, while the N-terminal carries the hypusine residue, which contains two positive charges and resembles spermidine, a molecule known to interact specifically with DNA and RNA. Indeed, hyp-eIF5A has been reported to bind to some RNA molecules in a sequence-specific manner
[28][29][28,29], and to assist with the transport of newly generated mRNAs from the nucleus to the cytoplasm
[28][30][28,30]. Moreover, eIF5A mutants exert a considerable impact on the balance between mRNA recruitment to ribosomes for translation and its degradation
[24][31][32][24,31,32], suggesting that eIF5A performs a function in the steps of mRNA decay downstream of decapping
[24][31][24,31]. Archaeal IF5A also plays a role in RNA metabolism as a moonlighting protein that associates with the ribosomes but also exerts RNAse activity
[33].
As previously stated, hyp-eIF5A assists in the translation of specific proteins that contain critical motifs in their amino acid sequences, although it is likely that we currently know only a small portion of its direct targets. Thus, the key role that eIF5A plays in different cellular processes is mostly due to the broad spectrum of cellular functions that its direct targets present. One of the major roles of hyp-eIF5A resides in cell proliferation and animal development. Eukaryotic translation initiation factor 5A and its hypusination are essential for cell proliferation in eukaryotes, and the disruption of eIF5A or DHPS genes, as well as the inhibitors of DHPS, cause growth arrest and strong anti-proliferative effects, including apoptosis
[13][34][35][36][37][38][39][40][41][13,34,35,36,37,38,39,40,41]. Hyp-eIF5A also mediates efficient autophagy through the translation of the autophagy master transcription factor TFEB and the ATG3 protein, the latter involved in the lipidation of LC3B and formation of the autophagosome
[42][43][42,43]. Additionally, eIF5A plays an important role in proper cytoskeleton organization and cell shape
[44][45][46][44,45,46] through the translation of formins in eukaryotes. In yeast, eIF5A is needed for the translation of the polyproline-containing formin Bni1, which is involved in polarized growth during mating
[47]. Accordingly, a mechanistic connection has been demonstrated between eIF5A and diaphanous, the formin involved in actin-rich cable assembly during the embryonic dorsal closure of
Drosophila and the migration of neural stem cells
[48]. Hyp-eIF5A has also been described to promote cell migration, invasion, and metastasis by controlling the expression of a set of key signalling molecules including RhoA and Rho-associated kinase, two cytoskeleton-regulatory proteins involved in promoting cell migration
[49], and by directly regulating MYC biosynthesis at specific pausing motifs
[50]. Specifically, EIF5A2 isoform has been shown to promote the epithelial-mesenchymal transition in several types of cancer cells
[51]. Eukaryotic translation initiation factor 5A has also been implicated in the regulation of apoptosis but the mechanism involved seems tangled given that this function appears to be opposite to the promotion of proliferation
[52][53][54][52,53,54]. It was recently found that, in response to stress, hyp-eIF5A promotes the translation of the tumor suppressor and pro-apoptotic factor p53, which contains polyproline motifs sensitive to the action of eIF5A
[55] and works as a transcription factor in charge of triggering a variety of antiproliferative programs.
The essential role eIF5A plays in the stated cellular processes implicates this protein in the pathogenesis of a wide variety of human diseases. Increasing evidence suggests that hyp-eIF5A plays an important role in modulating virus propagation. It has been defined as an essential cofactor of the human immunodeficiency virus type 1 (HIV-1) Rev transport factor. Through specific Rev binding, it participates in the translocation of unspliced viral mRNAs across the nuclear envelope
[56] and can behave as a nucleocytoplasmic shuttling protein
[57]. Although HIV was the first virus suggested to require eIF5A, this factor also participates in the replication of other viruses, such as the Marburg virus (MARV) and Ebola virus
[58]. A second human pathogenesis with a well-defined link to eIF5A is diabetes. In mouse models of diabetes, hyp-eIF5A in pancreatic islet β-cells is responsible for the translation of cytokine-induced transcripts, as well as for the activation and proliferation of T helper cells
[41][59][60][41,59,60].
The two paralogous genes encoding eIF5A,
EIF5A1 and
EIF5A2, are expressed under different conditions.
EIF5A1 is ubiquitously expressed in all mammalian tissues and cell types, whereas
EIF5A2 shows restricted expression in healthy tissue (being almost undetectable) but is overexpressed in certain tissues or cancer cells. The overexpression of both eIF5A isoforms has been observed in several tumors and triggers cell migration, invasion, and cancer metastasis (see review
[51] for details), but
EIF5A2 is considered a potential oncogene and diagnostic or prognostic marker
[61][62][61,62] because it is associated with poor survival, advanced disease stage, poor response to chemotherapeutic drugs, and metastasis. Genetic variants of eIF5A genes have been identified as the basis of certain rare neurodevelopmental disorders in humans
[63].
The inhibition of the eIF5A function has emerged as a potential target for treating the aforementioned diseases. The inhibition of eIF5A hypusination can be achieved by means of DHPS inhibitors, such as GC7 (N1-guanyl-1,7-diamonheptane), deoxyspergualin, or semapimod; DOHH inhibitors, such as ciclopirox, deferiprone, or mimosine; or inhibitors of ornithine decarboxylase (ODC), such as DFMO (difluoromethylornithine) (
Figure 1). DFMO
(di
fluoromethylornithine) is an irreversible inhibitor of ODC, which is the rate-limiting enzyme of polyamine biosynthesis. Therefore, DFMO acts to reduce polyamine levels and does not specifically inhibit eIF5A hypusination
[64]. DFMO has been used to decrease the replication of several RNA viruses, including Ebola, dengue, Zika, polio, and Coxsackievirus
[58][65][58,65] and in cancer prevention/therapy
[66]. Deferiprone and its structural analog, ciclopirox, are used in the treatment of iron overload and fungal infections, respectively. However, all three DOHH inhibitors affect the activity of other enzymes, such as proline hydroxylase enzyme
[37]. Among the known DHPS inhibitors, GC7, a diaminoheptane derivative, is the most efficient inhibitor (K
i value for GC7, 0.01 µM, compared to K
m for spermidine, 4.5 µM)
[67] and widely used today to inhibit eIF5A hypusination in mammalian cells
[68][69][68,69]. There are currently no inhibitors that act directly on eIF5A or, more selectively, on eIF5A2: this is a possible avenue for future research and development.
Lastly, the role of eIF5A in aging has been extensively studied in the last decade. eIF5A is implicated in long-term memory, adaptive immune response, cardiovascular function, and mitochondrial function; the failures of these processes are hallmarks of aging
[70].
2. Identification of Mitochondrial Processes and Targets under the Control of eIF5A
The main molecular role attributed to eIF5A is facilitating the translation of specific subsets of proteins containing eIF5A-dependent motifs
[10][11][10,11]. However, other less-characterized molecular roles have been proposed relating to the capacity of eIF5A to bind RNAs and regulate their metabolism. From the above results, it is clear that proper levels of hyp-eIF5A are necessary to preserve OXPHOS and mitochondrial function. The studies described above have proposed that eIF5A might be involved in the translation of specific mitochondrial proteins containing eIF5A-dependent MTS, in regulating the flux of metabolites to mitochondria to maintain aerobic metabolism through the regulation of glucose transporters, in sustaining mitochondrial dynamics through the interaction with proteins involved in mitochondrial fusion (Mnf1), in promoting autophagy by facilitating the synthesis of ATG3 and TFEB, or specifically in mitophagy through a mechanism that is dependent on PINK1 and Park proteins. To discern and prove these roles it will be necessary to identify the specific molecular process and/or the specific protein targets and conditions under the direct control of eIF5A.
In an attempt to identify mitochondrial protein candidates for direct eIF5A-dependent translation, we searched for peptide motifs causing eIF5A-dependent ribosome stalling
[11] in the
S. cerevisiae mitochondrial proteome. We found that eIF5A motif abundance in the 1117 nuclear-encoded mitochondrial proteins is only slightly higher than that in total yeast proteins, but these motifs are scarcely represented in the 10 mitochondrial DNA-encoded proteins (
Figure 2). Proteins involved in the TCA cycle show an average of 3.3 motifs/protein, higher than the average in the total yeast proteome (2.8 motifs/protein) and much higher than that in OXPHOS proteins (1.8 motifs/protein). However, the most represented eIF5A-dependent motifs were the same (GGA, GGG, and KPG) in TCA and OXPHOS proteins, with only two PPP motifs: one in the succinyl-CoA ligase (Lsc1) TCA enzyme, and another in the subunit of succinate dehydrogenase (Sdh4), which participates in the TCA cycle and OXPHOS.
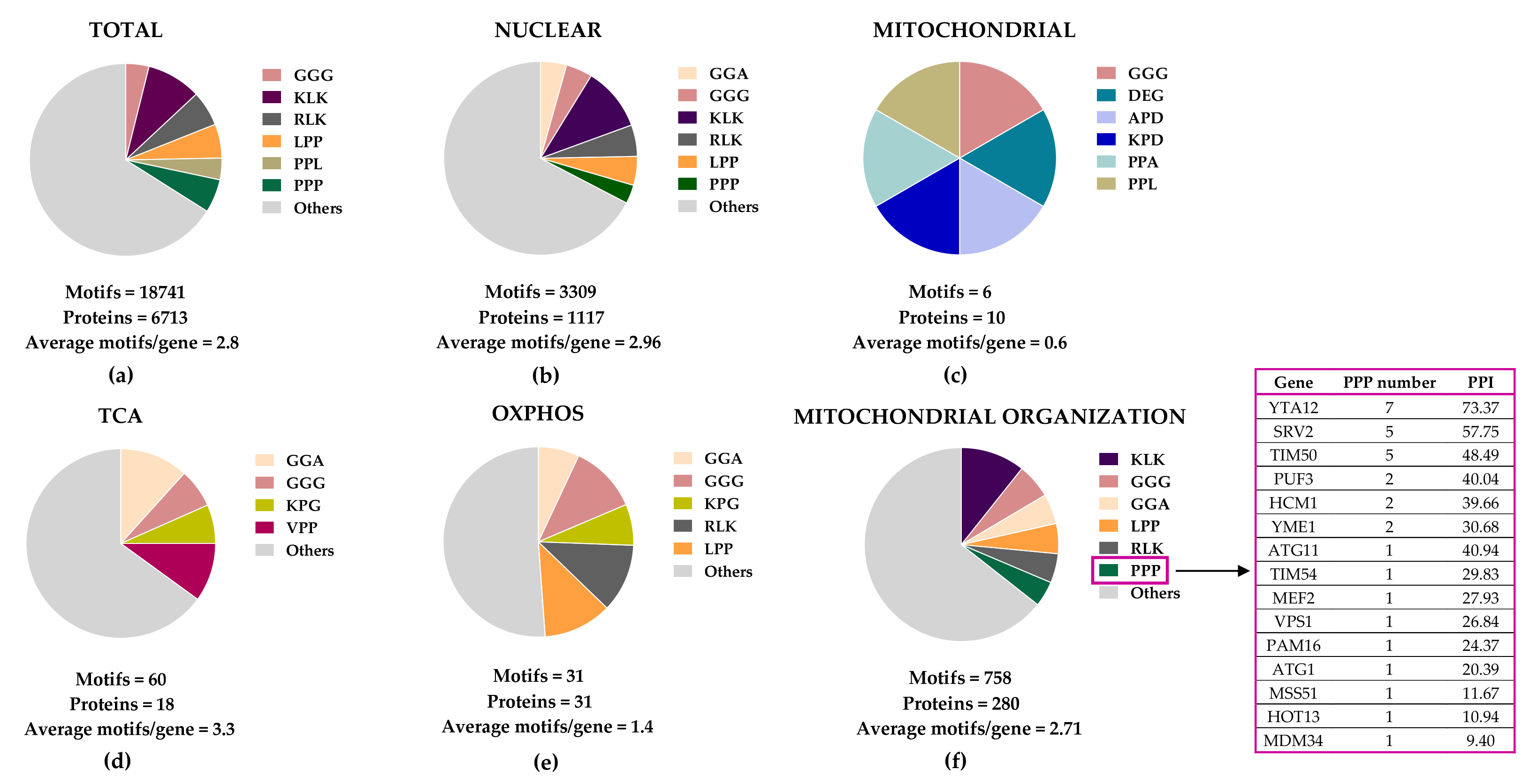 Figure 2.
Figure 2. Distribution of eIF5A-dependent motifs in
Saccharomyces cerevisiae mitochondrial proteins. Distribution of the 43 highest-scoring eIF5A-dependent ribosome-pausing tri-peptide motifs
[11] in the proteins of the whole yeast genome (
a), nuclear-encoded mitochondrial proteins (
b), mitochondrial-encoded proteins (
c), the tricarboxylic acid (TCA) cycle (
d), oxidative phosphorylation (OXPHOS) (
e), and in the mitochondrial organization Gene Ontology functional category (
f). The table shows the proteins involved in mitochondrial organization with at least one PPP motif. The protein pause index (PPI) is calculated as the sum of the quantitative value of the ribosome pause provoked by depletion of eIF5A in each of the 43 highest eIF5A-dependent tri-peptide motifs
[11] found in the amino acid sequence of each protein, and is higher in proteins that are putatively more dependent on eIF5A for their translation.
A search for yeast mitochondrial proteins with longer polyproline motifs showed that Yta12, Srv2, and Tim50 contain stretches of up to nine, six, and seven consecutive prolines, respectively (
Figure 2). Yta12 (homologous to human AFG3L2) is part of the conserved mitochondrial m-AAA protease, which is composed of the Afg3 and Yta12 proteins, and located at the inner mitochondrial membrane (
Figure 3). Yta12/Afg3 regulates mitochondrial proteostasis by mediating protein maturation and degradation and is required for the correct assembly of mitochondrial enzyme complexes
[71][166]. Interestingly, the Yta12/Afg3 complex is also involved in the splicing of the mitochondrial mRNAs containing introns
COX1 and
COB, which encode subunit 1 of cytochrome c oxidase and cytochrome b, respectively, which are part of the ETC. Thus, deficiency in Yta12/Yta10 causes low respiration due to, among other effects, deficient ETC assembly
[72][167]. Srv2 (homolog to human CAP1 and CAP2) mediates actin assembly at mitochondria, and deletion of Srv2 causes elongated hyperfused mitochondria and reduces respiration. Interestingly, Srv2 interacts with mitochondrial fission GTPase Dnm1/DRP1
[73][168]. Finally, Tim50 (a homolog of human TIMM50) is an essential subunit of the mitochondrial inner membrane TIM23 complex, which mediates the import of the majority of mitochondrial proteins through recognition of their MTSs (
Figure 3)
[74][169]. Although these three proteins are tempting candidates for mediating the effects of eIF5A on mitochondrial function, the presence of polyproline stretches is not a sufficient requirement to create dependence on eIF5A
[43][75][43,158]. Future studies will determine the precise mechanistic connections between eIF5A and the already described hyp-eIF5A-sensitive mitochondrial proteins and will identify new mitochondrial target proteins and processes under the control of eIF5A (
Figure 3).
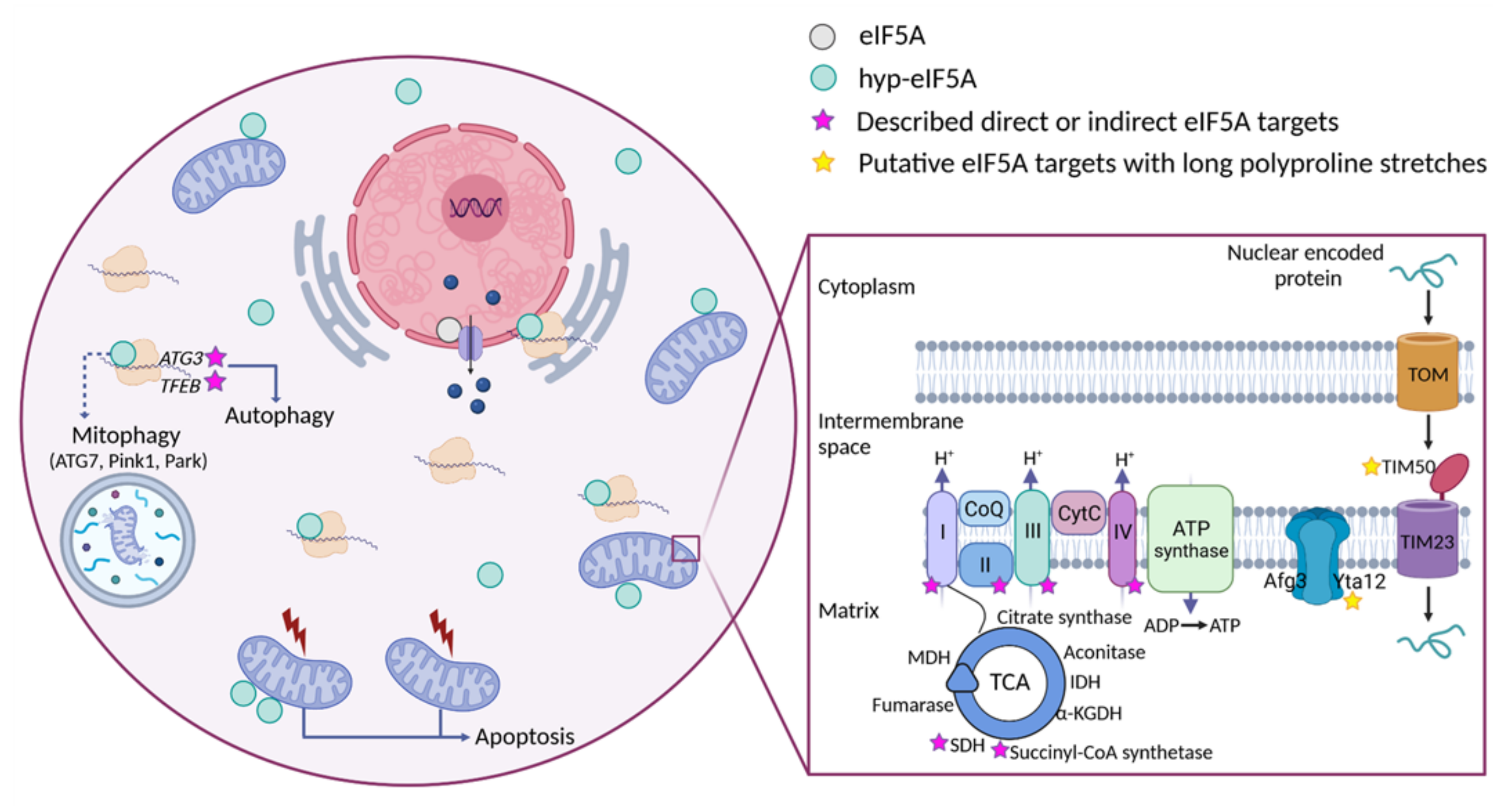 Figure 3.
Figure 3. Cellular funtions of eIF5A and model for its role in maintaining mitochondrial activity. eIF5A is known to be implicated in different cellular processes, although the most relevant and mitochondrial-related of these are represented in the Figure. Bound to ribosomes, hyp-eIF5A facilitates translation elongation at specific motifs
[10][11][10,11], as well as ER-coupled translation
[23][25][26][23,25,26]. In the nucleus, eIF5A helps to export certain mRNAs and proteins
[76][141]. eIF5A plays a controversial role in apoptosis, as it has been defined to be necessary to induce the mitochondrial mediated apoptosis
[77][78][124,146], but also to lead to cell death when inhibited
[79][133]. Hyp-eIF5A promotes autophagy through the translation of the autophagy factors ATG3 (autophagy-related 3) and TFEB (transcription factor EB)
[42][43][42,43]. Increasing evidence shows a direct link between hyp-eIF5A and mitochondrial function. In addition to its association with mitochondria
[80][81][82][78][143,144,145,146], some proteins of both the TCA and ETC have been described to be directly or indirectly affected under hyp-eIF5A inhibition
[83][84][126,147]. It has also been proposed that hyp-eIF5A could mediate mitophagy through ATG7 (Autophagy Related 7), Pink1 (the mitophagy-associated PTEN-induced putative kinase), and Park (the E3 ubiquitin ligase Parkin) proteins
[85][170]. Other proteins involved in the mitochondrial transport of nuclear-encoded proteins and mitochondrial organization are considered as putative eIF5A targets (
Figure 2). Among these, the mitochondrial inner-membrane integral proteins Yta12 (protease of the Yta12/Afg3 complex and yeast homolog of human AFG3L2), and Tim50 (essential subunit of the TIM23 complex and yeast homolog of human TIMM50) contain long polyproline stretches in their amino acids sequences, suggesting a possible dependence on eIF5A for their translation and, thus, a possible link between hyp-eIF5A and mitochondrial function. Figure processing was carried out using BioRender software.