The pathophysiology of pre-eclampsia involves two major pathways, namely systemic oxidative stress and subsequent generalised inflammatory response, which eventually culminates in endothelial cell injury and the syndrome of pre-eclampsia with multi-organ dysfunction. Aspirin has been used to reduce the risk of pre-eclampsia, but it only possesses anti-inflammatory properties without any antioxidant effect. Hence, it can only partially alleviate the problem. Tocotrienols are a unique form of vitamin E with strong antioxidant and anti-inflammatory properties that can be exploited as a preventive agent for pre-eclampsia. Many preclinical models showed that tocotrienol can also prevent hypertension and ischaemic/reperfusion injury, which are the two main features in pre-eclampsia.
1. Introduction
Pre-eclampsia is a clinical syndrome characterised by raised blood pressure and multi-organ dysfunction during the second half of pregnancy, labour or the puerperium in a previously normotensive woman [
1]. It is the leading cause of maternal and perinatal mortality and morbidity [
2]. The global incidence of pre-eclampsia has increased from 16.30 million in 1990 to 18.08 million in 2019 (a 17.80% increase) despite global improvement in maternal health [
3]. Chronic diseases, such as diabetes mellitus, obesity, chronic hypertension, renal disease, and connective tissue disorders, are risk factors of pre-eclampsia [
4].
Pre-eclampsia tends to recur, but a window of opportunity for its screening and prevention exists. The current management of pre-eclampsia involves screening (by risk factors, imaging, or biomarkers), counselling, continuous and meticulous monitoring and pharmacological control of blood pressure before and after delivery, as well as the prophylactic use of low-dose aspirin in high-risk women, magnesium sulphate to prevent eclampsia, and betamethasone to promote fetal lung maturity in pregnancies <34 weeks [
5]. Mapping the pathophysiology of a disease as accurately as possible is necessary to justify the use of the preventive measure and institute prevention by pharmacoprophylaxis. Over the years, many candidate drugs have been clinically tested, the most established being aspirin and calcium [
6]. Low-dose aspirin (81 mg/day) has been recommended by the US Preventive Task Force as the preventive drug, because it reduces pre-eclampsia risk and has multiple benefits on maternal and neonatal outcomes [
7]. However, a recent meta-analysis revealed that aspirin at doses lower than 150 mg/day could not remarkably prevent preterm pre-eclampsia [
8]. In addition, low-dose aspirin may be contraindicated in patients with hypersensitivity to non-steroidal anti-inflammatory drugs and gastrointestinal and genitourinary bleeding tendencies [
9]. Aspirin might also cross the placenta to the foetus and pose a theoretical risk of foetal intracranial haemorrhage despite the lack of epidemiological evidence [
10]. For calcium supplementation, the risk reduction is only about 20%, and is higher amongst populations with low calcium intake [
11]. Recent studies have demonstrated the potential of biologics, such as recombinant vascular endothelial growth factor (VEGF) and placental growth factor, in managing pre-eclampsia, but they are still in the development phase [
12]. Currently, the pursuit of an alternative agent for pre-eclampsia prevention is still ongoing.
Oxidative stress has been associated with pre-eclampsia in many observational studies [
13,
14]. Thus, many researchers have advocated for the potential role of antioxidants in preventing pre-eclampsia [
15]. The antioxidants that have been actively investigated include coenzyme Q10 [
16], melatonin [
17,
18], and curcumin [
19]. However, meta-analyses have reported that antioxidants are ineffective in pre-eclampsia prevention, and the effects on foetal outcomes are heterogeneous [
20,
21]. Results of trials involving vitamin E, particularly α-tocopherol, have been hugely disappointing [
22]. However, the effects of another form of vitamin E, tocotrienol, in pre-eclampsia prevention have been less explored. Only one study examined the effects of tocotrienol-rich fraction (TRF) derived from palm oil with promising outcomes [
23]. In this study, TRF reduced the risk of pre-eclampsia by 64% [
23].
2. Current Concept on the Pathophysiology of Pre-Eclampsia
The pathophysiology of pre-eclampsia has evolved progressively over the years, and the most recent concept embraces oxidative stress and the inflammatory process [
24]. The pathogenesis of pre-eclampsia involves maternal and placental pathways. The source of insult originates from maternal biochemical anomalies, due to chronic diseases or placental developmental anomalies in the form of restricted spiral artery invasion by foetal trophoblasts [
25]. Maternal origins of insult may arise from pre-existing diseases, such as diabetes mellitus, chronic hypertension, and obesity, whereas placental anomalies may be the result of immunological incompatibility between maternal and paternal human leukocyte antigens. Maternal metabolic diseases like ischaemia/reperfusion of the placental bed releases biochemical signals into the maternal circulation. These signals converge at the endothelium and form the final common pathway that results in pre-eclampsia, which is marked by endothelial dysfunction, raised blood pressure, and potential multi-organ complications. The humoral signals that produce pre-eclampsia and endothelial dysfunction consist of oxidative and inflammatory agents [
12,
25].
In the early stage of pregnancy, oxygen concentration within the trophoblast is maintained at a relatively low level to avoid teratogenic effects on the foetus. The hypoxic condition also up-regulates hypoxia-inducible factor 1 alpha, which, in turn, increases endothelial nitric oxide (NO) synthase to produce NO radicals that facilitate trophoblast proliferation and invasion. Towards the end of the first trimester, uteroplacental circulation is developed to allow a higher concentration of oxygen to reach the embryo to facilitate cell differentiation and foetal development. Oxidative stress develops easily at the syncytiotrophoblast, which is a special epithelium necessary for the transport of solute to the foetus and the production of hormones. It has a membrane rich in unsaturated fatty acid, but low in antioxidant enzymes; thus, it is susceptible to oxidative damage (summarised in [
26]).
Defective placental implantation onto the uterine wall impairs uterine spiral arteriole remodelling by extravillous trophoblasts, which subsequently reduces placental perfusion. The resultant cycles of placental ischaemia and reperfusion produce oxidative stress [
27]. Notably, the hypoxia/reperfusion cycles activate xanthine oxidase, nicotinamide adenine dinucleotide phosphate oxidase, and electron transport chain (complexes I and III) in the mitochondria to generate oxygen radicals. These radicals are reduced to hydrogen peroxide by manganese, copper, and zinc superoxide dismutase in the intermembranous space of the mitochondria, and subsequently reduced to water by glutathione peroxidase or catalase [
24]. Maternal metabolic derangements could result in impaired antioxidant defence and higher oxidants, which allow these radical oxygen species to cause cellular and tissue damage. Oxidative stress can also activate the nuclear factor kappa-B (NF-κB) pathway, which mediates the inflammation process [
24]. Damaged tissues release cell debris antiangiogenic factors, such as soluble fms-like tyrosine kinase-1 (sFlt-1), soluble endoglin, and cytokines [
27]. sFlt-1 binds to VEGF and reduces its bioavailability to maternal endothelial cells, which results in the reduction of NO production and triggers vasoconstriction [
28]. It also sensitises human umbilical artery endothelial cells to the action of pro-inflammatory cytokines [
29]. The hypoxic condition up-regulates p53 expression (indicative of pro-apoptotic signalling) and down-regulates Bcl-2 expression (indicative of anti-apoptotic signalling), which further escalates the apoptosis of syncytiotrophoblasts [
30,
31]. Oxidative stress also activates Wnt signalling and matrix metalloproteinases (MMPs), and thus promotes the invasiveness of trophoblasts and increases the risk of pre-eclampsia [
32].
Uterine immune cells are a major regulator of inflammation. During placentation, various immune cells, such as macrophages, natural killer cells, T-cells, B-cells, and dendritic cells, guide the proper invasion of trophoblasts [
33]. In pre-eclampsia, chronic placental and peripheral inflammation, marked by increased tumour necrosis factor-α (TNFα) and interleukin (IL)-6 and decreased IL-10 and IL-4, are observed [
34]. In particular, TNFα triggers endothelial dysfunction by reducing NO production, which activates the NF-κB pathway, propagates inflammation, increases endothelin-1 (ET-1), and causes vasoconstriction [
35]. TNFα, IL-6, and IL-17 also activate B-cells to secrete agonistic autoantibody against angiotensin II type-1 (AT-1) receptor, which, upon binding, causes the release of proinflammatory cytokines as well as ET-1 and sFlt-1 in the vasculature, and subsequently causes vasoconstriction [
36]. Overall, the pathophysiology of pre-eclampsia is summarised in
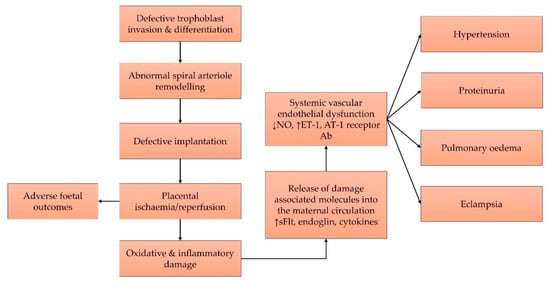
Figure 1.
Figure 1. Pre-eclampsia begins with defective placental implantation, which leads to recurrent ischaemia and reperfusion cycles that result in oxidative and inflammatory damages to the tissue. Damage-associated molecules cause systemic endothelial dysfunction and various features of pre-eclampsia. Abbreviations: Ab, antibody; AT-1, angiotensin-1; ET-1, endothelin-1; NO, nitric oxide; sFlt-1, soluble fms-like tyrosine kinase 1.
3. Vitamin E: Tocopherol and Tocotrienol
Although α-tocopherol was the first form of vitamin E to be recognised, natural vitamin E has eight lipophilic forms, including four tocopherols (α, β, γ and δ) and four tocotrienols (α, β, γ and δ) [
37]. Tocopherols are saturated forms of vitamin E, whereas tocotrienols are the unsaturated forms [
38]. α-Tocopherol is more ubiquitous in nature, whereas tocotrienol can be found in oil palm kernel, annatto bean, and rice bran. Tocotrienol isomers are usually found in mixtures of varying compositions in natural sources [
39]. For instance, palm-derived TRF consists of a mixture of ≈75% tocotrienol (α, β, γ and δ) and ≈25% α-tocopherol, whereas annatto tocotrienol consists of ≈90% δ-tocotrienol and ≈10% γ-tocotrienol [
40]. The effect of α-tocopherol on the treatment efficacy of tocotrienol mixtures is a subject of debate [
41], because α-tocopherol competes with other vitamin E isomers to bind to a-tocopherol transfer protein (ATTP) in the liver before being released into the circulation [
42]. α-Tocopherol preferentially binds with ATTP; thus, it has a higher bioavailability than other vitamin E isomers; the lower bioavailability might hinder the biological activities of the isomers. However, a comparative study did not find any remarkable difference in the skeletal and metabolic activities of palm TRF (containing α-tocopherol) and annatto tocotrienol (lacking α-tocopherol) in rats [
43,
44].
Vitamin E has been actively investigated for its effects in managing pre-eclampsia, because it could tackle several pathophysiological pathways of the disease, theoretically. Firstly, vitamin E is a well-established antioxidant [
45,
46] that can suppress the oxidative damage caused by repeated placental ischaemia/reperfusion. Secondly, vitamin E and its metabolites are anti-inflammatory agents [
47,
48,
49] that could suppress systemic inflammation caused by the release of damage-associated molecules. Thirdly, vitamin E supplementation may regulate blood pressure, but the evidence is debatable [
50].
Until recently, most studies have focused on α-tocopherol, which is the predominant form of vitamin E in body tissues. A previous Cochrane review demonstrated that vitamin E (α-tocopherol) combined with other supplements had no effect on pre-eclampsia [
51]. This result was because most women have sufficient vitamin E from their diet, and a short-term supplementation is unlikely to have an effect on pregnant women. In addition, most comparative studies showed that α-tocopherol is less efficacious than tocotrienols in managing chronic diseases [
40,
52,
53]. These limitations justify the shift of attention from α-tocopherol to tocotrienol in pre-eclampsia management.
4. Antioxidant Effects of Tocotrienol
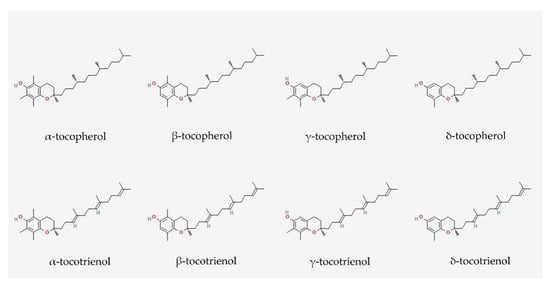
Tocopherols and tocotrienols share similar structures, such as an electron-rich chromanol ring and a long carbon tail. The carbon tail of tocopherols is saturated, whereas the tail of tocotrienols contains three double bonds (
Figure 2) [
54]. The structural difference between tocopherols and tocotrienols dictates their antioxidant potentials on a lipid membrane. Compared with α-tocopherol, α-tocotrienol has a stronger ability to disorganise lipid bilayers and is distributed more homogeneously within the lipid bilayers. α-Tocotrienol is also closer to the surface of the membrane, which facilitates its interaction with free radicals and the redox recycling process [
46,
55].
Figure 2. Molecular structures of tocopherols and tocotrienols. They consist of a chromanol ring and a long carbon tail. Tocopherols and tocotrienols have saturated and unsaturated carbon tails, respectively. Each isomer is distinct from each other by the position of the methyl side chain on the chromanol ring. The structures were obtained from PubChem.
Nuclear factor erythroid 2-related factor 2 (NRF2) is a master regulator of cellular antioxidants, as it governs the expression of various antioxidants in response to oxidative stress [
56]. Palm TRF up-regulates NRF2 expression, which increases the nuclear translocation of NRF2 in mouse liver [
57]. Moreover, δ-tocotrienol can stabilise the NRF2 activation induced by 5-fluorouracil in human oral keratinocytes, which can cause a sustained increase in the expression of heme oxygenase-1 and NAD(P)H:quinone oxidoreductase-1 [
58]. Another study on the antioxidant effects of δ-tocotrienol osteoblasts found that the activation of the NRF2 and PI3K/AKT pathways is necessary for the protection of δ-tocotrienol against oxidative stress [
59].
The antioxidant effects of tocotrienol, through direct electron scavenging activities or the activation of the NRF2 system, are translated in vivo. Tocotrienol isomers or mixtures exert antioxidant effects in various disease models, such as osteoporosis [
60,
61,
62], gastric ulcers [
63], cardiovascular diseases [
64,
65], and metabolic syndrome [
44]. Interestingly, a recent clinical trial demonstrated the more pronounced antioxidant benefits of TRF amongst women than men [
66]. A recent meta-analysis of human clinical trials showed that tocotrienol supplementation can improve the redox status of patients, as evidenced by the markedly reduced levels of lipid peroxidation products [
67]. Thus, the antioxidant effects of tocotrienol can be exploited to prevent the oxidative stress caused by placental ischaemia/reperfusion injury in pre-eclampsia.
5. Anti-Inflammatory Effects of Tocotrienol
The NF-κB pathway is an important regulatory pathway for inflammation [
68]. Previous studies showed that γ- and δ-tocotrienols inhibit the TNFα-induced activation of NF-κB by increasing intracellular dihydroceramides (a type of sphingolipid), cellular stress, and A20 [
69,
70]. A20 is a negative regulator of the NF-κB pathway by acting as a deubiquitinase [
71]. γ-Tocotrienol suppresses the phosphorylation and degradation of TNF-induced NF-κB inhibitor-α (IκBα) by preventing IκBα kinase activation and thus, abolishes the phosphorylation and nuclear translocation of p65 and the transcription of NF-κB-dependent reporter genes [
72]. This mechanism, together with the suppression of CCAAT/enhancer-binding protein beta, explains the suppressive effects of γ-tocotrienol on the expression of IL-6 and granulocyte colony-stimulating factor in primary bone marrow-derived macrophages, including chemokine response in the pathogenesis of asthma [
70].
NF-κB is also indispensable for the expression of cyclooxygenase, which is responsible for converting arachidonic acid to prostaglandin precursors [
73]. δ-Tocotrienol selectively decreases the expression of cyclo-oxygenase (COX)-2 and 5-lipoxygenase, but not COX-1, in lipopolysaccharide-induced microglial cells [
74]. This decrease is accompanied by a reduction in the expression of inducible NO synthase and IL-1β [
74]. The anticancer activities of γ-tocotrienol are also attributed to its ability to suppress the expression of COX-2, MMP-2, and MMP-9 proteins [
75]. A comparison of the anti-inflammatory effects of tocotrienol isomers and RAW264.7 macrophages showed that only α-tocotrienol can remarkably suppress TNFα expression [
76]. Besides, all tocotrienol isomers can reduce the expression of IL-6 and COX-2, but γ-tocotrienol is less efficacious in decreasing prostaglandin E
2 release. The COX-down-regulating activities of tocotrienol isomers are not shared by α-tocopherol [
76]. Another study on the anti-asthma potential of vitamin E reported that γ-tocotrienol is more effective than α-, γ-, and δ-tocopherols in suppressing IL-13-stimulated eotaxin-3 production in human lung epithelial A549 cells [
77]. These studies highlighted that the anti-inflammatory effects of each tocotrienol isomer against each cytokine could be different, and a mixture with the full spectrum of tocotrienols, such as palm TRF, could be more effective in suppressing inflammation.
The anti-inflammatory effects of tocotrienol mixtures can be explored with the aim of suppressing the inflammation caused by maternal metabolic conditions or inflammation in pre-eclampsia. Besides, animal studies also suggested that tocotrienols could prevent aspirin-induced gastric lesions by limiting lipid peroxidation [
78]. Tocopherols and tocotrienols have gastroprotective effects against oxidative stress, but only tocotrienols can block stress-induced changes in gastric acidity and gastrin level [
79]. Palm TRF increases the level of the constituently expressed COX-1 enzyme and prostaglandin E
2 to exert its gastric protective effects [
80]. γ-Tocopherol prevents, whereas α-tocopherol worsens, aspirin-induced gastric injury [
81]. Considering these studies, tocotrienols could mitigate the gastrointestinal bleeding side effects of aspirin. Nonetheless, a recent meta-analysis of human clinical trials showed that the effects of tocotrienol supplementation on circulating inflammatory mediators were not consistent [
67].