Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Shih-Hwa Chiou and Version 2 by Dean Liu.
Circular RNAs (circRNAs) are noncoding products of backsplicing of pre-mRNAs which have been established to possess potent biological functions. Due to their circular nature, the are characterized by high stability. Dysregulated circRNA expression has been linked to diseases including different types of cancer.
- circular RNA
- cancer progression
- metastasis
1. Circular RNAs (CircRNAs): Discovery, Biogenesis, and Degradation
1.1. Circular RNA Discovery
Originally, circular RNAs (circRNAs) were identified in eukaryotic cells in 1979 [1]. CircRNAs were initially considered as transcription noise devoid of any functionality. The advent of next-generation sequencing (NGS) in the early 2000s revealed the importance of noncoding transcriptome. CircRNAs were identified as noncoding RNA species with increased stability due to their circular nature, which fostered considerable investigation of their roles in the biological processes and disease [2][3][2,3].
1.2. CircRNA Biogenesis and Properties
CircRNAs are now known to be transcribed from protein-coding genes and further processed by unconventional pre-mRNA splicing mechanism, referred to as backsplicing, in which the 3′-end of an exon is ligated to the 5′-end donor splice site of the same or an upstream exon [3][4][3,4]. The conventional spliceosome and canonical splicing sites are essential for the process of backsplicing. Indeed, the process of backsplicing competes with conventional linear splicing [5]. Based on their structure derived from different modes of biogenesis, circRNAs can be classified as exonic (ecircRNAs), exon-intronic (EIciRNAs), circularized intronic (ciRNAs), and tRNA intronic (tricRNAs), with the former constituting 85% of all circRNAs. In the process of circRNA biogenesis, the distant donor and acceptor splice sites are selectively brought into proximity to be ligated, and three major models accounting for it have been delineated:
-
In a lariat-driven circularization, the exon-skipping linear splicing event produces a lariat that includes the skipped exons. Such lariat structure can further undergo internal splicing generating EIciRNAs or ecircRNAs (Figure 1A).
-
In intron pairing-driven circularization, also known as direct backsplicing, the splice sites are brought to close proximity by complementary pairing between inverted repeats (e.g., Alu repeats) flanking circularized exons (Figure 1B).
-
In RNA-binding protein (RBP)-driven circularization, the splice sites in intronic sequences are bridged together by certain trans-factor RBPs (Figure 1C).
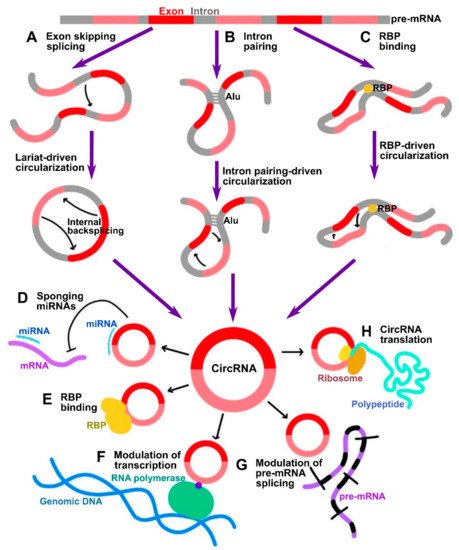
Figure 1. Summary of circRNA biogenesis and functional mechanisms. (A–C) Three major mechanisms of circRNA biogenesis by which distant backspliced sites are brought into proximity: (A) in lariat-driven circularization, circRNA is generated by internal backsplicing within a lariat formed by exon skipping direct splicing; (B) in intron pairing-driven circularization, backspliced sites are brought into proximity by pairing between inverted repeats such as Alu; (C) in RBP binding-driven circularization, backsplicing is facilitated by RBPs. (D–H) Summary of major biological functions of circRNAs: (D) miRNA sponging; (E) modulation of functions of RBPs; (F) modulation of transcription; (G) modulation of splicing; (H) translation of peptides.
Given the lower efficiency of backsplicing as compared to linear splicing, the production rate of circRNAs is lower than that of their linear counterparts. However, the lack of free ends renders them resistant to the exonuclease-mediated degradation which results in longer half-life and higher number within cells [6]. Based on this property, circRNAs can be used to serve as novel diagnostics markers or disease targets. In 2019, it was first reported that circRNAs undergo biodegradation by RNase L global RNA-degrading endonuclease [7]. In such a mechanism, circRNAs were found to be stabilized by binding to inactive unphosphorylated protein kinase R (PKR) that blocked circRNAs’ accessibility to RNase L [7][8][7,8]. Phosphorylation of PKR activates it and mediates its dissociation from circRNAs, which, in turn, exposes them to active RNase L [7] (Figure 2). Similarly, another study showed that the highly structured nature of circRNAs ensures their degradation via binding to RNA-binding protein UPF1 and its associated protein G3BP1, which causes subsequent degradation by RNase P or RNase MRP [9]. Therefore, reswearchers can conclude that the cell achieves circRNA homeostasis by possessing a well-controlled circRNA biogenesis and degradation systems (Figure 2). Dysregulation of these processes can lead to cellular aberrations and disease. Numerous transcriptome analysis studies identified more than 20,000 circRNAs expressed from 60% of total genes in eukaryote cells. Intriguingly, a portion of circRNAs is expressed in a tissue- or developmental stage-specific manner indicative of temporal and spatial control of their expression. CircRNAs have been identified in numerous species by different high-throughput RNA deep sequencing projects. In one of the studies, it was identified that around 14.4% of human fibroblast circRNAs can be linked to 69 murine testis orthologous circRNAs [10]. Such orthologous conservation features of circRNA imply the importance of their role in gene regulation.
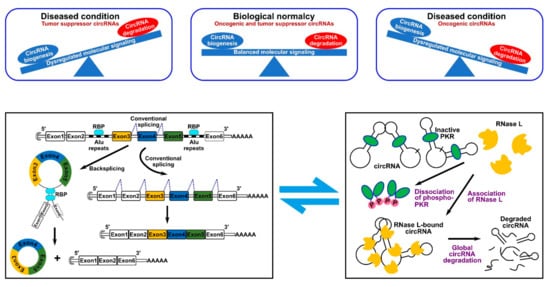
Figure 2. CircRNA homeostasis determines normal biological conditions or disease state. (Top panel) such homeostasis is determined by the fine balance between circRNA biogenesis and degradation mechanisms. (Bottom panel) schematic representation of a typical RBP-driven biogenesis mechanism (left) and of the RNase L-dependent degradation mechanism (right).
2. Molecular Functions of CircRNAs
CircRNAs have been proven to be involved in various biological processes and mechanistically have been widely characterized to conduct their functions by several major mechanisms:
-
One of the most investigated functions of circRNAs is their ability to indirectly control gene expression through sponging of microRNAs (miRNAs) [6] (Figure 1D). CircRNAs often contain miRNA response elements (MREs) which can bind miRNAs, thus preventing them from silencing their gene targets [11][12][11,12]. By such mechanism, these circRNAs are classified as competing endogenous RNAs (ceRNAs) [11]. Among the most prominent of them is ciRS-7 (CDR1as), which was discovered to possess up to 70 target sites for miR-7 [6]. Therefore, a vast number of studies has been focused on investigating ceRNA mode of action of circRNAs. However, large-scale screening studies indicate that usually circRNAs contain a limited number of MREs, indicative that this is not the only bona fide mechanism.
-
CircRNAs can associate with RNA binding proteins (RBPs) and thus mediate different aspects of their functionality (Figure 1E). For instance, circRNAs can act as scaffolds facilitating the formation of protein complexes. For example, circ-FOXO3 was shown to interact with both MDM2 and p53, which promoted MDM2-dependent ubiquitination and degradation of p53 [13]. The same circRNA was also shown to form ternary complex with CDK2 and p21, preventing the association of the former into functional complexes with cyclins A and E, thus acting by decoy mechanism [14].
-
CircRNAs that retain the intronic sequences of their parental genes, such as ciRNAs and EIciRNAs, are predominantly localized in the nucleus. As was shown by Pol II CLIP, EIciRNAs such as circEIF3J and circPAIP2 could associate with RNA polymerase II and enhance their parental gene expression in cis in a U1 snRNP-dependent manner [15] (Figure 1F). The broad-scale effects of circRNAs on pre-mRNA splicing are not widely characterized as yet; however, it has been known that circularization and linear splicing compete with each other on a co-transcriptional level (Figure 1G). Such regulatory mechanism was demonstrated for MBL/MBNL1 gene encoding muscleblind (MBL) splicing factor. The protein product of this gene was shown to directly promote circularization of the exon 2 of its host gene by direct binding to the flanking sequences in the adjacent introns to produce circMBL, concomitantly reducing linear splicing [5]. Finally, in addition to regulating mRNAs at transcriptional and splicing levels, circRNAs were also shown to regulate the stability of mRNAs. For example, the degraded fragments of CDR1, as circRNA could downregulate CDR1 by promoting degradation via antisense pairing [16]. On the contrary, circRasGEF1B could enhance the stability of its target ICAM1 mRNA [17].
-
Although circRNAs have traditionally been regarded as noncoding RNAs given the lack of the 5′ cap that is normally required for the initiation of translation, the discovery of internal ribosome entry sites (IRES) within multiple circRNAs suggests that they may be extensively translated into peptides or proteins in 5′ cap-independent manner [18] (Figure 1H). One prominent example of such circRNAs is circ-ZNF609, whose IRES-dependent translation was shown to play an important role in myogenesis [19]. In addition, it was shown that initiation of translation in circRNAs can be driven by m6A methylation. CircRNAs were found to be enriched in consensus m6A motifs, whose methylation by METTL3/14 methyl transferase complex promoted the translation, and demethylation by FTO demethylase inhibited the translation [20]. In such a mechanism, m6A deposited on circRNAs could be recognized by YTHDF3 m6A reader protein, which in turn recruited eIF4G2 initiation factor [20].