Cold-stored platelets are making a comeback. They were abandoned in the late 1960s in favor of room-temperature stored platelets due to the need for longer post-transfusion platelet recoverability and survivability in patients with chronic thrombocytopenia. However, the current needs for platelet transfusions are rapidly changing. Today, more platelets are given to patients who are actively bleeding, such as ones receiving cardiac surgeries. It has been established that cold-stored platelets are more hemostatically effective, have reduced bacterial growth, and have longer potential shelf lives. These compelling characteristics led to the recent interest in bringing back cold-stored platelets to the blood systems. However, before reinstating cold-stored platelets in the clinics again, a thorough investigation of in vitro storage characteristics and in vivo transfusion effects is required.
- cold-stored platelets
- product storage
- storage solution
- platelet transfusion
1. Introduction
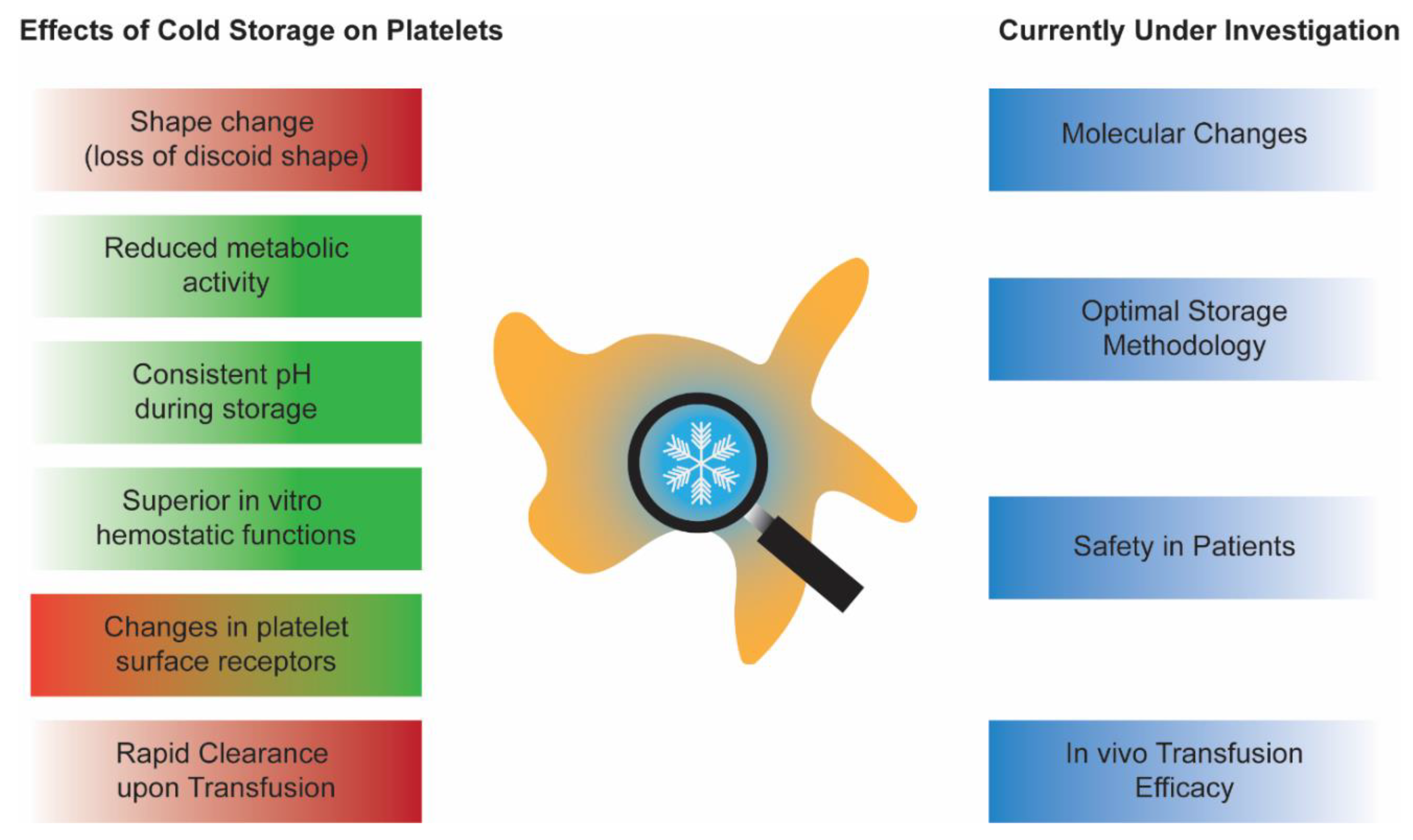
2. CPs In Vitro Characterization: An Update and a Debate
3. “Omics” and Cold-Stored Platelets
4. Reduction of CPs Storage Lesion, a Work in Progress
Results from the recent omics study showed that oxidative stress played a significant role in platelet storage lesion both in RPs and CPs. Hence, to improve CPs’ storage quality, both synthetic and naturally occurring antioxidants have been used as supplements. N-acetylcysteine (NAC) has been used as a supplement to minimize the mitochondrial uncoupling and proton leaks caused by cold-storage [32]. NAC supplement also showed the slight effect of reducing ROS in CPs. Both RPs and CPs with NAC supplement had better correction in a mice tail bleeding experiment than CPs without NAC [32]. However, these experiments had relatively few biological replicates. In addition, the use of platelet additive solution (PAS) could have influenced the results as well. In another study, whole blood-derived platelet rich plasma (PRP) supplemented with NAC and stored in the cold seemed to have reduced platelet activation measured by P-selectin expression compared to RPs [33]. NAC supplement did not result in differences in the phosphatidylserine expression of CPs or RPs. However, the platelets used in this experiment were stored in gas-permeable tubes instead of standardized storage containers, making it difficult to draw solid conclusions [33]. In a follow-up study, the effect of NAC on CPs post-transfusion recovery has been tested using the Prkdcscid mouse model [34]. There was a slight improvement of platelet recovery 2 h post transfusion in the CPs with the NAC group. However, this improved recovery effect was diminished after 24 h post transfusion [34]. It is important to note that in this mouse study, CPs without NAC supplement were not tested due to aggregate formation in the storage bag. The absence of this control makes data interpretation more challenging, as the result could be due to either the storage temperatures or the addition of NAC. Another antioxidant, resveratrol, and a mitochondrial targeting protein, cytochrome c, have been used to reduce oxidative stress in CPs as well [35]. The preliminary results from these supplements showed decreased ROS in resveratrol-treated CPs [35]. However, further investigation of supplemented CPs function and transfusion efficacy are needed.
In addition to using antioxidants, synthetic molecules have also been used to reduce the lesion in CPs. Specialized pro-resolving mediators (SPMs) are biosynthesized molecules derived from essential fatty acids. SPMs have been found to reduce acute inflammation during the host response to invading pathogens [36]. Specific SPMs, such as resolving E1, have been found to regulate platelet activations [37]. In CPs, the addition of SPMs significantly reduced the glycoprotein 1b expression compared to CPs without SPMs [38]. However, in this study, there were no RPs with SPMs as a control. It is not clear what other effects SPMs may have in CPs during storage. Similarly, sodium octanoate (SO) was used as a supplement in CPs to improve mitochondrial activity and reduce CPs apoptosis [39]. Although CPs with SO seem to trend toward better mitochondrial activity compared to CPs without SO, the results were not statistically significant. Furthermore, although a lower phosphatidylserine expression was observed with CPs with SO on day 5 of storage, further analysis of the caspase-3 level between the treatment groups was not significant. Platelet endocytosis with HepG2 cell models also did not show a difference in SO-treated versus non-treated CPs [39]. Finally, integrin inhibitor RGDS and a calcium ion chelator EGTA were added to improve cold-stored platelet post-transfusion clearance [40]. It is important to note that this experiment was performed with mouse platelets, and the cold storage period of murine platelets was for a maximum of 24 h. While the calcium chelator EGTA showed some improvement in cold-stored murine platelet clearance, the effect disappeared when EGTA was used in human platelets. In addition, EGTA supplementation in human PRP prevented the reduction in platelet count commonly observed in CPs during storage as well as aggregate formation. However, CPs stored in PAS could also achieve similar effects, so the exact function of EGTA in CPs is not as straightforward as it seems.5. Safer CPs: Pathogen Inactivation and Platelet Additive Solutions
Despite stringent donor eligibility criteria, blood products are still exposed to the risk of emerging pathogen contaminations. West Nile outbreaks in the past and the recent outbreak of Zika virus in the US are good examples of this [41][42]. Pathogen inactivation (PI) technology has been attracting interest in further securing the safety of blood components [43]. This technology targets and damages the nucleic acids of pathogens using ultraviolet light, thus preventing pathogen replication in blood products. PI RPs have been licensed in many countries including the US, Spain, and Switzerland, according to a report by AABB [44]. CPs, due to the lower storage temperature, have been shown to slow the growth of bacteria [12]. However, psychrophilic bacteria, such as Yersinia enterocolitica, can survive at 4 °C and proliferate to a clinically significant level [45]. Yersinia enterocolitica has been associated with transfusion reaction in red blood cell transfusions [46]. Thus, there are incentives in investigating the effect of PI on CPs. Recent data show that CPs in PAS treated with Intercept PI (Cerus Corporation, Concord, CA, USA) had less activation level and aggregation in response to agonists than non-PI treated CPs [47]. However, when tested in microfluidic devices, PI-treated CPs had a better fibrin formation rate [47]. One caveat for this study is that the platelets were only stored for up to 5 days in the cold. When Intercept PI-treated CPs were stored for an extended period up to 21 days, there was no major difference in the metabolism of the platelets in terms of count, pH, glucose, or lactate compared to non-PI-treated CPs [48]. Interestingly, PI treatment resulted in significant changes to CPs function. There was a significant reduction in clot retraction capabilities in PI-treated CPs. There was also elevated phalloidin staining in PI-treated CPs. Furthermore, there was also an elevated lysis index at 30 min measured by ROTEM in PI-treated CPs [48]. These data suggest that PI may adversely affect CPs during storage. A different PI system, Theraflex UV-platelet System (UVC), was also used on CPs [49]. Similar to the effects of the Intercept system, this technology also resulted in damage to the CPs, as shown by decreased HSR response. In addition, PI-treated CPs also had higher activation shown by annexin V staining and higher release of microparticles compared to untreated CPs [49]. Together, these results show that PI does have significant effects on the cold storage of platelets. More data on the post-transfusion effectiveness of PI-treated CPs are needed to fully understand the feasibility of these platelets in the clinic. Platelet additive solution (PAS) is a chemical solution that can replace plasma in platelet storage. Different types of PAS used in studying CPs have been summarized in Table 1. PAS provides several benefits. First, PAS can be used to replace some of the plasma in platelet storage. Then, the replaced plasma can be used for fractionation into intravenous immunoglobulin (IVIg) to help with the ongoing shortages [50][51]. In addition, reducing the amount of plasma in a platelet unit lowers the risk of allergic transfusion reactions [52]. This adds an additional layer of safety for the transfusion recipients. In CPs, there have been suggestions that PAS could be beneficial [53]. It was first established that similarly to PAS in RPs, a residual of 35% plasma is optimal for CPs in stored PAS [54]. Based on this result, the effects of different PAS on CPs storage quality have been investigated. For example, two FDA-approved collections and PAS storage systems of platelets (Trima apheresis collection system in Isoplate PAS versus Amicus collection system in Intersol PAS) stored in the cold were compared [55]. The results showed that there were only minor differences in the platelet count over 15 days of storage between the two systems. CPs produced from the two systems also had similar aggregation response profiles [55]. It is important to note that the minor differences observed could be contributed by either the different apheresis collection instruments or by the different PAS. To further differentiate the storage quality of CPs in PAS versus in plasma, ISO PAS platelets were stored for 19 days and compared to apheresis platelets in 100% plasma [56]. CPs stored in PAS had significantly reduced glucose consumptions versus CPs in plasma. This is likely due to the substitution of acetate as an energy source instead of glucose, while also reducing lactate production. Day 19 CPs PAS also had acceptable pH, which would meet FDA pH criteria [56]. One criticism for this work is that the CPs in 100% plasma were only stored for 3 days, making parallel comparison to day 19 CPs in PAS difficult. CPs stored in PAS-E in also maintained pH above 6.4 for 21 days [57]. The aggregation response to agonists is the same between CPs in PAS versus CPs in plasma. However, it is important to note that the addition of PAS to CPs reduced fibrinogen concentration and other coagulation factors, which could be detrimental to the recipient upon transfusion. In addition, PAS could provide additional benefits specific to CPs. The lowered fibrinogen concentration in PAS leads to a significant reduction of aggregate formation while preserving the in vitro function in CPs [10]. Additionally, PAS may help in maintaining mitochondrial integrity by reducing the depolarization of the inner membrane potential in CPs [58]. Finally, the type of PAS used in CPs could also affect the post-transfusion recovery. The recoveries of CPs stored in either 100% plasma, Intersol, or Isoplate PAS were compared in an in vivo transfusion of autologous radiolabeled platelets into healthy donors. Interestingly, CPs stored in Intersol had better recoveries than CPs in Isoplate. However, CPs stored in 100% plasma still had better than CPs stored in either PAS [59].| PAS | Author | Ref | Main Usage |
|---|---|---|---|
| T-PAS | Nair et al. | [21] | Investigation of the clot structure of CPs. |
| T-PAS | Reddoch-Cardenas et al. | [56] | Compared CPs stored in PAS versus RPs and CPs in plasma. |
| T-PAS and Intersol | Getz et al. | [10] | Reduce aggregate formation in CPs using PAS. |
| SSP+ | Six et al. | [47] | Pathogen inactivation (Intercept). |
| SSP+ | Johnson et al. | [49] | Pathogen inactivation (Theraflex UV-platelet System). |
| SSP+ | Marini et al. | [54] | Plasma replacement and test for the percentage of residual plasma required. |
| SSP+ | Wood et al. | [60] | Delayed cold storage. |
| PAS-E | Hegde et al. | [32] | NAC supplementation in CPs |
| PAS-E | Johnson et al. | [57] | Prolonged platelet cold storage. |
| Intersol | Agey et al. | [48] | Pathogen inactivation (Intercept). |
| Isoplate | Reddoch-Cardenas et al. | [58] | Preserving mitochondrial function in CPs. |
| Isoplate and Intersol | Reddoch-Cardenas et al. | [55] | Compare the CPs storage characteristics in two FDA approved PAS. |
| Isoplate and Intersol | Stolla et al. | [59] | PAS in CPs in vivo recoveries using autologous radiolabeled platelet transfusion. |
| PAS-IIIM | Braathen et al. | [61] | Delayed cold storage. |
References
- Murphy, S.; Gardner, F.H. Effect of storage temperature on maintenance of platelet viability—Deleterious effect of refrigerated storage. N. Engl. J. Med. 1969, 280, 1094–1098.
- Josefsson, E.C.; Gebhard, H.H.; Stossel, T.P.; Hartwig, J.H.; Hoffmeister, K.M. The macrophage alphaMbeta2 integrin alphaM lectin domain mediates the phagocytosis of chilled platelets. J. Biol. Chem. 2005, 280, 18025–18032.
- Badlou, B.A.; Spierenburg, G.; Ulrichts, H.; Deckmyn, H.; Smid, W.M.; Akkerman, J.-W.N. Role of glycoprotein Ib? In phagocytosis of platelets by macrophages. Transfusion 2006, 46, 2090–2099.
- Rumjantseva, V.; Grewal, P.K.; Wandall, H.H.; Josefsson, E.C.; Sørensen, A.L.; Larson, G.; Marth, J.D.; Hartwig, J.H.; Hoffmeister, K.M. Dual roles for hepatic lectin receptors in the clearance of chilled platelets. Nat. Med. 2009, 15, 1273–1280.
- Wandall, H.H.; Hoffmeister, K.M.; Sørensen, A.L.; Rumjantseva, V.; Clausen, H.; Hartwig, J.H.; Slichter, S.J. Galactosylation does not prevent the rapid clearance of long-term, 4 °C-stored platelets. Blood 2008, 111, 3249–3256.
- Zucker, M.B.; Borrelli, J. Reversible alterations in platelet morphology produced by anticoagulants and by cold. Blood 1954, 9, 602–608.
- White, J.G.; Krivit, W. An ultrastructural basis for the shape changes induced in platelets by chilling. Blood 1967, 30, 625–635.
- Reddoch, K.M.; Pidcoke, H.F.; Montgomery, R.K.; Fedyk, C.G.; Aden, J.K.; Ramasubramanian, A.K.; Cap, A.P. Hemostatic function of apheresis platelets stored at 4 °C and 22 °C. Shock 2014, 41, 54–61.
- Oliver, A.E.; Tablin, F.; Walker, N.J.; Crowe, J.H. The internal calcium concentration of human platelets increases during chilling. Biochim. Biophys. Acta (BBA)-Biomembr. 1999, 1416, 349–360.
- Getz, T.M.; Montgomery, R.K.; Bynum, J.A.; Aden, J.K.; Pidcoke, H.F.; Cap, A.P. Storage of platelets at 4 °C in platelet additive solutions prevents aggregate formation and preserves platelet functional responses. Transfusion 2016, 56, 1320–1328.
- Bynum, J.A.; Meledeo, M.; Getz, T.M.; Rodriguez, A.C.; Aden, J.K.; Cap, A.P.; Pidcoke, H.F. Bioenergetic profiling of platelet mitochondria during storage: 4 °C storage extends platelet mitochondrial function and viability. Transfusion 2016, 56, S76–S84.
- Ketter, P.M.; Kamucheka, R.; Arulanandam, B.; Akers, K.; Cap, A.P. Platelet enhancement of bacterial growth during room temperature storage: Mitigation through refrigeration. Transfusion 2019, 59, 1479–1489.
- Becker, G.A.; Tuccelli, M.; Kunicki, T.; Chalos, M.K.; Aster, R.H. Studies of platelet concentrates stored at 22 C and 4 C. Transfusion 1973, 13, 61–68.
- Zhao, H.W.; Serrano, K.; Stefanoni, D.; D’Alessandro, A.; Devine, D.V. In Vitro Characterization and Metabolomic Analysis of Cold-Stored Platelets. J. Proteome Res. 2021, 20, 2251–2265.
- Sharma, V.; Shah, M.; Pullela, R.; Cohen, A.J. Platelet Utilization in a Tertiary Care Hospital: An Opportunity for Reduced Transfusion? Blood 2010, 116, 2575.
- Pandey, S.; Belanger, G.A.; Rajbhandary, S.; Cohn, C.S.; Benjamin, R.J.; Bracey, A.W.; Katz, L.M.; Menitove, J.E.; Mintz, P.D.; Gammon, R.R. A survey of US hospitals on platelet inventory management, transfusion practice, and platelet availability. Transfusion 2021, 61, 2611–2620.
- Reddoch-Cardenas, K.; Bynum, J.; Meledeo, M.; Nair, P.; Wu, X.; Darlington, D.; Ramasubramanian, A.; Cap, A. Cold-stored platelets: A product with function optimized for hemorrhage control. Transfus. Apher. Sci. 2018, 58, 16–22.
- Berzuini, A.; Spreafico, M.; Prati, D. One size doesn’t fit all: Should we reconsider the introduction of cold-stored platelets in blood bank inventories? F1000Research 2017, 6, 95.
- Getz, T.M. Physiology of cold-stored platelets. Transfus. Apher. Sci. 2019, 58, 12–15.
- Nair, P.M.; Pandya, S.G.; Dallo, S.F.; Reddoch, K.M.; Montgomery, R.K.; Pidcoke, H.F.; Cap, A.P.; Ramasubramanian, A.K. Platelets stored at 4 °C contribute to superior clot properties compared to current standard-of-care through fibrin-crosslinking. Br. J. Haematol. 2017, 178, 119–129.
- Nair, P.M.; Meledeo, M.A.; Wells, A.R.; Wu, X.; Bynum, J.A.; Leung, K.P.; Liu, B.; Cheeniyil, A.; Ramasubramanian, A.K.; Weisel, J.W.; et al. Cold-stored platelets have better preserved contractile function in comparison with room temperature-stored platelets over 21 days. Transfusion 2021, 61, S68–S79.
- Melchinger, H.; Jain, K.; Tyagi, T.; Hwa, J. Role of Platelet Mitochondria: Life in a Nucleus-Free Zone. Front. Cardiovasc. Med. 2019, 6, 153.
- Koessler, J.; Klingler, P.; Niklaus, M.; Weber, K.; Koessler, A.; Boeck, M.; Kobsar, A. The Impact of Cold Storage on Adenosine Diphosphate-Mediated Platelet Responsiveness. TH Open 2020, 4, e163–e172.
- Miles, J.; Bailey, S.L.; Obenaus, A.M.; Mollica, M.Y.; Usaneerungrueng, C.; Byrne, D.A.; Fang, L.; Flynn, J.R.; Corson, J.; Osborne, B.; et al. Storage temperature determines platelet GPVI levels and function in mice and humans. Blood Adv. 2021, 5, 3839–3849.
- Scorer, T.G.; FitzGibbon, L.; Aungraheeta, R.; Sharma, U.; Peltier, G.C.; McIntosh, C.S.; Reddoch-Cardenas, K.M.; Meyer, A.; Cap, A.P.; Mumford, A.D. TEG PlateletMapping assay results may be misleading in the presence of cold stored platelets. Transfusion 2020, 60, S119–S123.
- Gutmann, C.; Joshi, A.; Mayr, M. Platelet “-omics” in health and cardiovascular disease. Atherosclerosis 2020, 307, 87–96.
- D’Alessandro, A.; Thomas, K.A.; Stefanoni, D.; Gamboni, F.; Shea, S.M.; Reisz, J.A.; Spinella, P.C. Metabolic phenotypes of standard and cold-stored platelets. Transfusion 2019, 60, S96–S106.
- Wang, S.; Jiang, T.; Fan, Y.; Zhao, S. A proteomic approach reveals the variation in human platelet protein composition after storage at different temperatures. Platelets 2018, 30, 403–412.
- Nagy, M.; Mastenbroek, T.G.; Mattheij, N.J.A.; De Witt, S.; Clemetson, K.J.; Kirschner, J.; Schulz, A.S.; Vraetz, T.; Speckmann, C.; Braun, A.; et al. Variable impairment of platelet functions in patients with severe, genetically linked immune deficiencies. Haematologica 2017, 103, 540–549.
- Hickey, M.J.; Hagen, F.S.; Yagi, M.; Roth, G.J. Human platelet glycoprotein V: Characterization of the polypeptide and the related Ib-V-IX receptor system of adhesive, leucine-rich glycoproteins. Proc. Natl. Acad. Sci. USA 1993, 90, 8327–8331.
- Mukai, N.; Nakayama, Y.; Ishi, S.; Murakami, T.; Ogawa, S.; Kageyama, K.; Murakami, S.; Sasada, Y.; Yoshioka, J.; Nakajima, Y. Cold storage conditions modify microRNA expressions for platelet transfusion. PLoS ONE 2019, 14, e0218797.
- Hegde, S.; Wellendorf, A.M.; Zheng, Y.; Cancelas, J.A. Antioxidant prevents clearance of hemostatically competent platelets after long-term cold storage. Transfusion 2020, 61, 557–567.
- Handigund, M.; Bae, T.W.; Lee, J.; Cho, Y.G. Evaluation of in vitro storage characteristics of cold stored platelet concentrates with N acetylcysteine (NAC). Transfus. Apher. Sci. 2016, 54, 127–138.
- Handigund, M.; Kim, J.T.; Bae, T.W.; Lee, J.; Cho, Y.G. N-acetylcysteine reduce the stress induced by cold storage of platelets: A potential way to extend shelf life of platelets. Transfus. Apher. Sci. 2020, 60, 103039.
- Ekaney, M.L.; Grable, M.A.; Powers, W.F.; McKillop, I.H.; Evans, S.L. Cytochrome c and resveratrol preserve platelet function during cold storage. J. Trauma Acute Care Surg. 2017, 83, 271–277.
- Basil, M.C.; Levy, B.D. Specialized pro-resolving mediators: Endogenous regulators of infection and inflammation. Nat. Rev. Immunol. 2015, 16, 51–67.
- Fredman, G.; Van Dyke, T.E.; Serhan, C.N. Resolvin E1 Regulates Adenosine Diphosphate Activation of Human Platelets. Arter. Thromb. Vasc. Biol. 2010, 30, 2005–2013.
- Reddoch-Cardenas, K.M.; Sharma, U.; Salgado, C.L.; Cantu, C.; Darlington, D.N.; Pidcoke, H.F.; Bynum, J.A.; Cap, A.P. Use of Specialized Pro-Resolving Mediators to Alleviate Cold Platelet Storage Lesion. Transfusion 2020, 60, S112–S118.
- Baghdadi, V.; Yari, F.; Nikougoftar, M.; Rafiee, M.H. Platelets Apoptosis and Clearance in The Presence of Sodium Octanoate during Storage of Platelet Concentrate at 4 °C. Cell J. 2020, 22, 212–217.
- Xiang, B.; Zhang, G.; Zhang, Y.; Wu, C.; Joshi, S.; Morris, A.J.; Ware, J.; Smyth, S.S.; Whiteheart, S.W.; Li, Z. Calcium Ion Chelation Preserves Platelet Function During Cold Storage. Arter. Thromb. Vasc. Biol. 2020, 41, 234–249.
- Lanteri, M.C.; Kleinman, S.H.; Glynn, S.A.; Musso, D.; Hoots, W.K.; Custer, B.S.; Sabino, E.C.; Busch, M.P. Zika virus: A new threat to the safety of the blood supply with worldwide impact and implications. Transfusion 2016, 56, 1907–1914.
- Harrington, T.; Kuehnert, M.J.; Kamel, H.; Lanciotti, R.S.; Hand, S.; Currier, M.; Chamberland, M.E.; Petersen, L.R.; Marfin, A.A. West Nile virus infection transmitted by blood transfusion. Transfusion 2003, 43, 1018–1022.
- Seltsam, A. Pathogen Inactivation of Cellular Blood Products—An Additional Safety Layer in Transfusion Medicine. Front. Med. 2017, 4, 219.
- Listing of Countries in Which Pathogen Inactivation Technology Systems and Products Are in Use. 2015 March. Available online: https://www.aabb.org/docs/default-source/default-document-library/regulatory/eid/prt-systems-in-use-country-listing.pdf (accessed on 24 December 2021).
- Ribault, S.; Faucon, A.; Grave, L.; Nannini, P.; Faure, I.B. Detection of Bacteria in Red Blood Cell Concentrates by the Scansystem Method. J. Clin. Microbiol. 2005, 43, 2251–2255.
- Guinet, F.; Carniel, E.; Leclercq, A. Transfusion-Transmitted Yersinia enterocolitica Sepsis. Clin. Infect. Dis. 2011, 53, 583–591.
- Six, K.R.; Devloo, R.; Compernolle, V.; Feys, H.B. Impact of cold storage on platelets treated with Intercept pathogen inactivation. Transfusion 2019, 59, 2662–2671.
- Agey, A.; Reddoch-Cardenas, K.; McIntosh, C.; Sharma, U.; Cantu, C.; Cap, A.; Bynum, J. Effects of Intercept pathogen reduction treatment on extended cold storage of apheresis platelets. Transfusion 2020, 61, 167–177.
- Johnson, L.; Cameron, M.; Waters, L.; Padula, M.P.; Marks, D.C. The impact of refrigerated storage of UVC pathogen inactivated platelet concentrates on in vitro platelet quality parameters. Vox Sang. 2018, 114, 47–56.
- Bayry, J.; Kazatchkine, M.D.; Kaveri, S.V. Shortage of human intravenous immunoglobulin—reasons and possible solutions. Nat. Clin. Pr. Neurol. 2007, 3, 120–121.
- N’Kaoua, E.; Attarian, S.; Delmont, E.; Campana-Salort, E.; Verschueren, A.; Grapperon, A.-M.; Mestivier, E.; Roche, M. Immunoglobulin shortage: Practice modifications and clinical outcomes in a reference centre. Rev. Neurol. 2021, S0035-3787(21)00768-2.
- Mathur, A.; Swamy, N.; Thapa, S.; Chakraborthy, S.; Jagannathan, L. Adding to platelet safety and life: Platelet additive solutions. Asian J. Transfus. Sci. 2018, 12, 136–140.
- Van der Meer, P.F.; de Korte, D. Platelet Additive Solutions: A Review of the Latest Developments and Their Clinical Implications. Transfus. Med. Hemotherapy 2018, 45, 98–102.
- Marini, I.; Aurich, K.; Jouni, R.; Nowak-Harnau, S.; Hartwich, O.; Greinacher, A.; Thiele, T.; Bakchoul, T. Cold storage of platelets in additive solution: The impact of residual plasma in apheresis platelet concentrates. Haematologica 2019, 104, 207–214.
- Reddoch-Cardenas, K.M.; Montgomery, R.K.; Lafleur, C.B.; Peltier, G.C.; Bynum, J.A.; Cap, A.P. Cold storage of platelets in platelet additive solution: An in vitro comparison of two Food and Drug Administration–approved collection and storage systems. Transfusion 2018, 58, 1682–1688.
- Reddoch-Cardenas, K.M.; Sharma, U.; Salgado, C.L.; Montgomery, R.K.; Cantu, C.; Cingoz, N.; Bryant, R.; Darlington, D.N.; Pidcoke, H.F.; Kamucheka, R.M.; et al. An in vitro pilot study of apheresis platelets collected on Trima Accel system and stored in T-PAS+ solution at refrigeration temperature (1–6 °C). Transfusion 2019, 59, 1789–1798.
- Johnson, L.; Vekariya, S.; Wood, B.; Tan, S.; Roan, C.; Marks, D.C. Refrigeration of apheresis platelets in platelet additive solution (PAS-E) supports in vitro platelet quality to maximize the shelf-life. Transfusion 2021, 61, S58–S67.
- Reddoch-Cardenas, K.M.; Peltier, G.C.; Chance, T.C.; Nair, P.M.; Meledeo, M.A.; Ramasubramanian, A.K.; Cap, A.P.; Bynum, J.A. Cold storage of platelets in platelet additive solution maintains mitochondrial integrity by limiting initiation of apoptosis-mediated pathways. Transfusion 2020, 61, 178–190.
- Stolla, M.; Fitzpatrick, L.; Gettinger, I.; Bailey, S.L.; Pellham, E.; Christoffel, T.; Slichter, S.J. In vivo viability of extended 4 °C-stored autologous apheresis platelets. Transfusion 2018, 58, 2407–2413.
- Wood, B.; Johnson, L.; Hyland, R.A.; Marks, D.C. Maximising platelet availability by delaying cold storage. Vox Sang. 2018, 113, 403–411.
- Braathen, H.; Sivertsen, J.; Lunde, T.H.F.; Kristoffersen, E.K.; Assmus, J.; Hervig, T.A.; Strandenes, G.; Apelseth, T.O. In vitro quality and platelet function of cold and delayed cold storage of apheresis platelet concentrates in platelet additive solution for 21 days. Transfusion 2019, 59, 2652–2661.