1. Introduction
Liver pathology still accounts a significant portion in the structure of morbidity
[1][2][1,2]. In this regard, a constant search for new therapeutic approaches, including the usage of cellular and gene technologies, is needed
[3][4][3,4]. Resident macrophages of the liver, Kupffer cells, which account 80–90% of all macrophages of the mammalian organism
[5], are considered as one of the possible application targets of regenerative medicine
[6]. In addition, Kupffer cells play a key role in maintaining liver homeostasis in normal and pathological conditions
[7]. Macrophages serve as significant regulators of liver regeneration
[8][9][8,9], development of inflammatory processes, fibrosis
[10][11][10,11], cholestatic diseases
[12], tumor progression
[13], and liver damage during transplantation
[14].
Development of new methods of treating liver diseases is impossible without studying the population of liver macrophages. However, this is an extremely difficult task, as macrophages represent a highly heterogeneous cell population. In this regard, the review focuses on current data on ontogenesis, phenotypic and functional plasticity of liver macrophages and their role in reparative processes.
2. Macrophage Population of the Liver
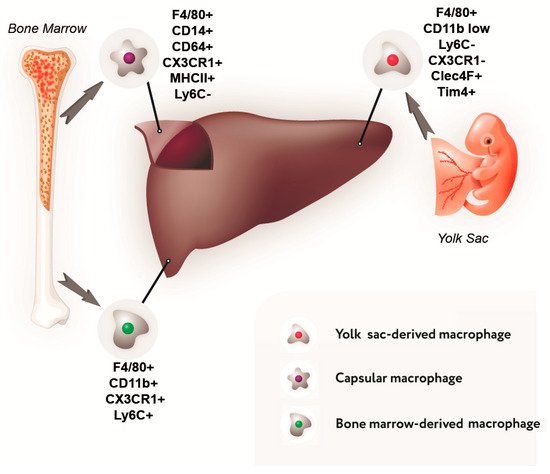
As has been mentioned above, in the normal liver of mammals, the population of macrophages is composed largely of cells that originate from the erythro-myeloid progenitor cells of the yolk sac and are commonly referred to as Kupffer cells. Macrophages derived from the blood monocytes represent a minor population in the liver; according to different estimates, they constitute from 5% to 30% of total liver macrophages
[15][16][17][18][15,23,24,25].
Such a large variation in quantitative estimation of the population of bone marrow-derived macrophages in the liver is due to specific choices of the markers used to identify them
[19][26]. In our opinion, the most relevant choice is represented by a combination of Ly6C and CX3CR1 markers
[15][20][18][15,17,25].
The majority of studies on macrophage ontogenesis were performed using laboratory mice. The postnatal mouse macrophages of bone marrow origin carry the surface protein Ly6C, which is absent from the surface of Kupffer cells
[18][21][25,27]. Similarly, the protein marker CX3CR1 is used to recognize bone marrow-derived macrophages in rats (since Ly6C protein is not detected in rats)
[18][25]. According to the data obtained with these markers, the proportion of bone marrow-derived macrophages in the liver constitutes approximately 5%
[15][20][18][15,17,25]. Such an assessment is consistent with the observation that liver macrophages are diverse by their size and location. Two types of liver macrophages have been identified: large, located near the sinusoidal capillaries, and small, located around the central veins and portal tracts. Both of them express CD68; however, CD163 is expressed at a high level only by large Kupffer cells
[22][23][28,29]. The small macrophages constitute about 8%, which is similar to the proportion of bone marrow-derived liver macrophages determined by Ly6C immunostaining
[15][18][15,25]. These findings allow the possibility to consider small liver macrophages as the cells of monocytic origin.
The most controversial data is obtained using markers CD11b and CD68. These markers are expressed in a variety of cell types including all leukocytes; in addition, CD68 is found in endothelial cells and fibroblasts
[24][30]. CD11b and CD68 participate in cell adhesion, cell migration, and phagocytosis; accordingly, their expression may undergo considerable rapid changes
[24][25][26][30,31,32].
CD11b has been employed as a marker of bone marrow-derived macrophages in some studies
[16][17][27][23,24,33]. It has been shown that, although all macrophages found in the liver (Kupffer cells and bone marrow-derived macrophages) express CD11b, the bone marrow-derived macrophages express it at a higher level
[20][28][18][29][30][17,19,25,34,35].
Using the latest Single Cell RNA sequencing method in the human liver MacParland and colleagues established the presence of two populations of resident macrophages with pro-inflammatory and immunoregulatory (non-inflammatory) phenotypes. One of the main differences between these two populations was the expression level of the MARCO marker, which prevailed in non-inflammatory macrophages; MARCO+ macrophages were localized in the periportal zone
[31][36]. Another study with Single cell RNA sequencing in a mouse steatohepatitis model also revealed two macrophage populations: Kupffer cells expressing Clec4f and monocytes-derived macrophages expressing Lyz2
[32][37]. Moreover, the population of macrophages with monocytic origin, found in the liver with steatohepatitis, was also extremely heterogeneous. The authors distinguish at least three subtypes of such macrophages: MoMF I showed a high expression of the extracellular matrix protein fibronectin 1 (Fn1), as well as microsomal glutathione S-transferase 1 (Mgst1) and methionine sulfoxide reductase B1 (Msrb1). MoMF II expressed few marker genes including Chil1, mainly indicating a bone marrow origin. MoMF III was characterized by a pronounced expression of Il1b
[32][37].
In summary, according to one set of reports, the resident mouse liver macrophages have Ly6C
−CX3CR1
low phenotype, whereas under the inflammatory conditions the liver tissue becomes infiltrated by the bone marrow-derived macrophages with Ly6C
+CX3CR1
hi phenotype
[18][33][34][25,38,39]. Other studies indicate that Kupffer cells are either F4/80
+CD11b
−CD169
+CD68
+CD80
lo, or CD68
+CD11b
−, or F4/80
highCD11b
low, whereas the infiltrating bone marrow-derived monocytic macrophages are F4/80
+CD11b
+CD80
hi CD11b
+ [16][29][30][35][23,34,35,40]. The data available for other mammalian species are limited. For humans, it has been proposed to use CD163L as a Kupffer cell-specific marker and CLEC5A as a marker specific for the monocyte-derived pro-inflammatory macrophages migrating to the liver
[36][41].
Data on the composition of the liver macrophage population and their phenotypic characteristics are summarized in Figure 1.
Figure 1. Sources of development and immunophenotype of a subpopulation of liver macrophages.
3. The Diversity of Macrophage Functional Types
The macrophages represent heterogeneous cell population not only by their developmental sources, but also by their functional characteristics. Macrophages are capable of very rapid phenotypic and functional changes under the action of signal molecules.
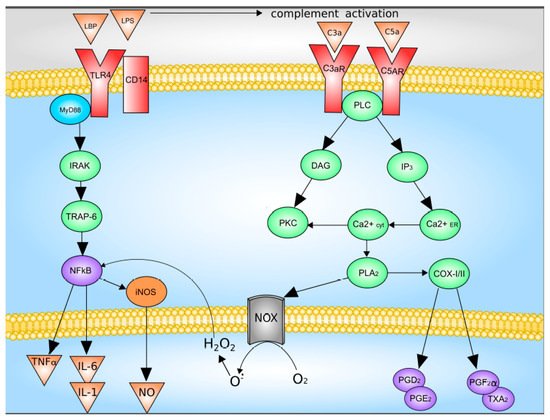
The main activators of Kupffer cells are complement factors C3a and C5a, as well as LPS
[37][38][47,48]. LPS directly through TLR4 activates Kupffer cells, which leads to an increase in the synthesis and release of TNF-a, IL-1b, IL-6, IL-12, IL-18, IL-10, and IFN-γ. The binding of LPS and TLR4 leads to the formation of a multiprotein membrane complex containing myeloid differentiation factor MyD88, TNFR-associated factors (TRAFs), interleukin-1 receptor-associated kinases (IRAKs), and TGF-beta-activated kinase 1 (TAK1). The formation of this complex leads to the activation of JNK and p38 mitogen-activated protein kinases (MAPKs) and NF-kB signaling pathway were activated
[5][37][38][5,47,48].
Upon activation of the complement system, C3a and C5a bind to their receptors with followed activation of down-stream signaling system which includes G proteins and phospholipase C. This pathway leads to increased synthesis of Kupffer cells Prostaglandin D2, Prostaglandin E2, Prostaglandin F2α, as well as thromboxane A2, as well as the release of superoxide
[5][39][5,49]. Data on the Kupffer cells activation are summarized in
Figure 2.
Figure 2. Mechanisms of Kupffer cell activation. C3 and C5, complement factor 3 and 5; C3a and C5a, activated complement factor 3 and 5; CD14, CD14 receptor; COX-I/II, cyclooxygenase-I/II; DAG, diacylglycerol; H
2O
2, hydrogen peroxide; iNOS, inducible nitric oxide synthase; IL-1, interleukin-1; IL-6, interleukin-6; IP3, inositol-3-phosphate; IRAK, interleukin-1 receptor-associated kinase; LPB, LPS-binding protein; LPS, lipopolysaccharide; NOX, NADPH oxidase; NFκB, nuclear factor κB; NO, nitric oxide; O
2−, superoxide anion; PGD2, PGE2 and PGF2α, Prostaglandin D2, Prostaglandin E2, Prostaglandin F2α; PKC, protein kinase C; PLA2, phospholipase A2; PLC, phospholipase C; TLR4, Toll-like receptor 4; TNFα, tumor necrosis factor-α; TRAP-6, TNF-activated factor 6; TXA2, thromboxane A2. With modification made in NetworkPainter from
[5] under CC-BY.
Another way to activate Kupffer cells is through the formation of inflammasomes—a complex of NOD-like receptors and apoptosis-associated speck-like protein a CARD, which is necessary for activation of caspase 1, which in turn activates pro-inflammatory cytokines IL-1β and IL-18
[40][41][50,51]. Formation of inflammasomes in the cytosol plays a central role in the development of inflammatory and fibrotic diseases. The most known for the development of liver diseases is the NLRP3 inflammasome, their increased formation leads to severe inflammation in the liver, which is associated with overproduction of IL-1β and severe liver neutrophilic infiltration
[41][51].
In situ activation of macrophages (the so-called ‘macrophage polarization’) may take either the M1 pro-inflammatory direction or the M2 anti-inflammatory direction
[37][42][47,52]. The M1 and M2 polar states of macrophage activation differ not only by specific markers, but, most importantly, by their roles in immune response and tissue repair
[43][53]. A local shift towards M2 phenotype within a tissue damage area significantly improves the dynamics and efficiency of repair processes, as has been demonstrated for a number of particular models including skin wound healing
[44][54], spinal cord damage
[45][55], myocardial infarction and cardiomyopathy
[46][56] and some other models. A shift in the balance of different functional classes of macrophages may occur by suppression of the M1 activation pathway as well as by reinforcement of the M2 activation pathway. At the same time, a part of macrophages may remain in the non-activated state
[37][47][47,57]. In vivo shifts in the M1/M2 balance may be induced in a paracrine manner e.g., by blocking IL6 which promotes the M1 polarization
[45][55] or introducing IL4
[48][58] or IL10 which promote the M2a and M2c polarization, respectively
[49][59].
However, the use of markers is in many respects conditional. No exact correspondence exists between the Ly6C+ and M1 macrophages as well as between Ly6C- and M2 macrophages. On the contrary, Ly6C+ macrophages may in some cases give rise to both M1 and M2 functional types
[50][51][60,61].
An experimental study by our research group has revealed that macrophages of bone-marrow origin and Kupffer cells have similar profiles of surface markers. Both of them express CD86 which is necessary for activation, proliferation, cytokine production and T cell differentiation
[52][62], CD163 which is involved in bacterial recognition and triggering of local inflammatory reactions
[53][63] and CD206 which is required for phagocytosis, antigen presentation and clearance of pro-inflammatory mediators from the circulation
[54][55][64,65]. However, some quantitative differences in the expression of surface markers are evident: monocytic macrophages predominantly express the M1 marker CD86, whereas Kupffer cells predominantly express the M2 markers CD163 and CD206. At the same time, despite the association of surface markers with specific functionalities, the increased expression levels have been observed in both types of macrophages under the influence of bacterial endotoxin lipopolysaccharide (LPS) combined with IFN-γ or IL4 combined with IL10. Similar results have been obtained in the analysis of cytokine gene expression: transcription of pro-inflammatory cytokine genes (
Il1b,
Il6,
Tnfa) and anti-inflammatory cytokine genes (
Il10) in macrophages under the influence of LPS combined with IFN-γ or IL4 combined with IL10 becomes upregulated in concert
[56][66].
To summarize, no strict border between M1 and M2 functional types of macrophages can be drawn. Instead, there is a continuous series of transitional forms being constantly updated to the needs of particular tissue microenvironments
[38][57][58][48,67,68].