3. Occurrence of BRs in Plants
BRs have been found in organs and all parts of the plants, such as leaves, stems, roots, flowers, pollen, anthers, and seeds
[17][18][17,18]. BR is universally distributed in all growing tissues of higher plants, but significantly higher concentrations have been detected in seed, pollen, and fruit
[19]. The level of BR in the young tissues (1–100 ng/g fresh weight) is normally higher than in mature ones (0.01–0.1 ng/g fresh weight)
[20]. BL and CS are the most significant BRs because of their higher biological activity and widespread distribution in plants. However, due to its commercial availability, 24-epibrassinolide (EBR) is the most commonly used BR for studying the physiological effects of exogenous steroid phytohormones on plants
[21].
After the BL discovery, about 69 BRs have been identified in 64 plant species, including 6 gymnosperms, 53 angiosperms (41 dicotyledons and 12 monocotyledons), 1 pteridophyte (
Equisetum arvense), 3 algae (
Hydrodictyon reticulatum, Cystoseira myrica, and
Chlorella Vulgaris) and 1 bryophyte (
Marchantia polymorpha)
[17][18][17,18], a moss (
Physcomitrella patens), lycophytes (
Selaginella moellendorffii and
S. uncinata), and 13 fern species
[22]. The concentration of 6-deoxotyphasterol (6-deoxoTY) concentration was found to be 6400-fold greater than BL in the pollen of
Cupressus arizonica. Additionally, the highest concentration of BR, 6.4 mg 6-deoxoTY per kilogram (kg) pollen, has been found in
C. arizonica [17]. Only 52 BRs have been reported in terms of their biological activities in plants
[16]. The CS, BL, TY, 6-deoxoCS, TE, and 28-norCS are the most abundant BRs in plants
[14][15][20][14,15,20]. The most extensive variety of BRs (2 conjugated and 25 free forms) was found in unripe bean (
Phaseolus vulgaris) seeds
[20]. Other legumes having fewer BR members have been found in shoots, seeds, and pollen, and their quantity was between 0.007 and 628l g/g fresh weight
[14].
4. Chemical Structure of BRs
BRs have been categorized into three major types on the basis of each steroid molecule’s carbon number (i.e., C
27, C
28, and C
29)
[23]. The 5α-cholestane skeleton is the basic structure of C
27-BRs, 5α-ergostane for C
28-BRs, while 5α-stigmastane is the basic structure of C
29-BRs. The structure of these hormones differs because of the type and orientation of oxygenated functions of A and B rings, and the number and position of functional groups in the side chain of the molecule. These changes occur as the result of oxidation and reduction reactions during biosynthesis. In general, BRs have been classified into conjugated (5) free, and (64) compounds
[23].
BRs have vicinal hydroxyl groups in relation to the A-ring at C-2α and C-3α. BRs with α and β-hydroxyls, or ketone at the C-3 position are the progenitors of the BRs with 2α, 3α-vicinal hydroxyls. BR containing 2α, 3β-, 2β, 3α-, or 2β, 3β-vicinal hydroxyls maybe the precursors of 2α, 3α-vicinal hydroxyls. Two 2α, 3α-vicinal hydroxyl groups on the A-ring are responsible for overall structural attributes of the most active BRs, such as BL and CS. The declining order of activity 2α, 3α > 2α, 3β > 2β, 3α > 2β, 3β suggests that the α-oriented hydroxyl group at C-2 is essential for biological activity of BRs in plants
[24][25][24,25]. On the basis of cholestane side chain, BRs have been divided by different substituents into C-23, C-24, and C-25: 23-oxo (4 compounds), 24-methylene (3 compounds), 24
S-ethyl (4 compounds), 24
R-methyl (5 compounds), 24
S-methyl (23 compounds), C-25, 24-ethylidene (3 compounds), 24-methyl-25-methyl (3 compounds), 24-methylene-25-methyl (6 compounds) without substituent at C-23 (3 compounds), without substituent at C-24 (8 compounds), and without substituents at C-23, C-24 (2 compounds)
[26][27][28][26,27,28].
BRs are classified into 6-oxo (6-ketone) (34 compounds), and 6-deoxo (non-oxidized) (21 compounds), and 7-oxalactone (12 compounds) types depending on B-ring oxidation stage. However, only one fourth type BR with hydroxyl group at C-6, such as 6α-hydroxycastasterone (6α-OH-CS) has been observed. In contrast, two compounds, for instance, 28-nor-(22
S)-22-hydroxycampestrol (28-nor-22-OHCR) and (22
S)-22-hydroxycampestrol (22-OHCR) were identified as a fifth type of BRs. Generally, 7-oxalactone BRs have been observed to show stronger biological activity compared to 6-deoxo type, and 6-oxo type. Sometimes 6-oxo BRs show activity similar to 7-oxalactone compounds, while non-oxidized BRs exhibit essentially little activity in the bean internode test or very little in the rice lamina inclination test
[29][30][31][29,30,31].
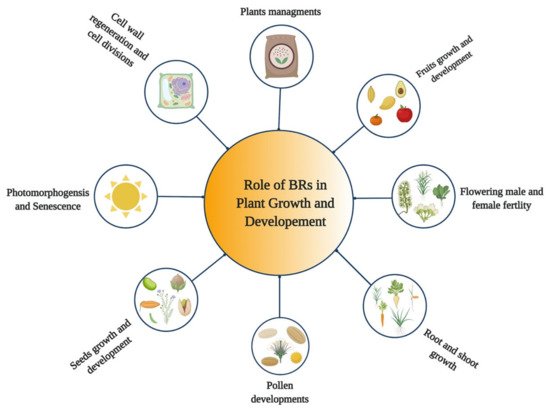
5. Role of BRs in Growth and Development of Plants
BRs are steroid hormones that play various roles in the growth and development of plants
[32][33][34][32,33,34] (
Figure 1). BRs regulate various developmental and physiological processes in plants, such as expansion, cell division, stem cell maintenance, vascular development, elongation of different cell types, and floral transition
[35][36][37][38][35,36,37,38]. Moreover, they play diverse roles in hypocotyl elongation
[39], root growth
[7][40][7,40], shoot growth
[28], stomata patterning
[41][42][41,42], pollen tube growth, seed germination, and pollen germination and development
[43], treachery element differentiation
[44], xylem formation
[45][46][45,46], xylem differentiation, photomorphogenesis and plant reproduction
[47][48][47,48], and senescence
[49]. BRs have the ability to activate the cell cycle during seed germination
[50], regulate cell cycle progression
[51], control leaf cells’ proliferation
[52], and induce excessive growth in hydroponically grown plants
[53]. BRs also regulate the abiotic and biotic stress responses and stomata development
[7][34][54][7,34,54]. Moreover, BR plays a vital role in regulating male and female fertility in crops
[55][56][55,56]. BR plays a role in etiolation and promotes the elongation of stigma
[19], plant architecture, thermo-tolerance, proton transport, tiller number, leaf angle, and leaf size
[57][58][57,58]. In addition, the exogenous application of BR or alteration in their biosynthesis and signaling could improve crop yields
[59][60][59,60].
Figure 1.
Role of Brassinosteroids (BRs) in growth and development of plants.
BRs are also involved in the regulation of several genes in plants (
Table 1). In a study, Chen et al.
[61] found that BRs induced
WRKY46,
WRKY54, and
WRKY70 genes that were observed to play positive roles in BR-regulated plant growth in
A. thaliana. In another study, the histone lysine methyltransferase SDG8 is involved in BR-regulated gene expression. The knockout mutant
sdg8 displayed a reduced growth phenotype with compromised BR responses in
A. thaliana [62]. In
A. thaliana, BR regulates the seed development and affects the seed size/weight and number by transcriptionally modulating the genes and pathways that regulate the development of the seed and ovule
[63][64][63,64]. Furthermore, BRs regulate root Nitrogen foraging response in
A. thaliana during mild Nitrogen deficiency. A Brassinosteroid signaling kinase 3 (
BSK3) gene is involved in the elongation of primary root during mild Nitrogen deficiency
[65].
BR regulates the grain yield and plant architecture in rice
[23]. The gene products of
BRD1 and
D11 are involved in the biosynthesis of BR, and affect the height of rice plants
[66]. The
OsDwarf2/OsDwarf1 reduction encodes a C-6 oxidase needed for BR biosynthesis in rice, reducing second internode and seed length elongation
[66][67][66,67]. In rice, BRs mediate the effects of N fertilization on spikelet development and contribute to promote spikelet growth by increasing the level of antioxidant system (AOS) and energy charge during panicle development
[68]. In maize, inadequate BR biosynthesis causes male sterility due to failure of pollen and anther growth
[55][56][55,56]. In cotton, both the fiber initiation and elongation of cultured cotton ovules have been reported to require BRs
[69][70][69,70]. Moreover, an exogenous application of EBR delays the vegetative to generative transition in monocotyledonous wheat (
Triticum aestivum L.). Brassinazole, a BR inhibitor, accelerates the transition and heading stage
[71]. The application of BR accelerated winter rapeseed maturation by 4 to 8 days
[72].
Priming of lucerne seeds with BL considerably increased length, vigor, and fresh and dry shoot and root weights
[73]. The application of BR promoted the ripening of grape berry
[74]. Treatment with EBR was observed to substantially increase sucrose synthase (sucrolytic) and soluble sugars content in berries
[75]. Treatment with BR helped in reducing the decay of jujube fruits, likely due to its ability to postpone senescence and induce disease resistance
[76]. Additionally, in potato tubers, the BRs have been shown to promote apical meristem growth
[77], accelerating the cell division rate in isolated protoplasts of
Petunia hybrida [78]. In
Pharbitis nil, CS and BL treatments inhibit flowering in combination with the inductive photoperiod, implying that BR works in tandem with environmental cues to ensure the proper reproductive transition
[19][79][19,79].
BRs can participate in physiological processes in response to stress by tuning plant growth, and improving plant performance by interacting with plant growth regulators or other plant hormones
[80][81][80,81]. The disruption of BR signaling affects several developmental processes, including seed development
[64], pollen development
[82], and flowering time
[83]. Plants with BR deficiency are dwarfed
[84], and exhibit altered stomatal development
[41][85][41,85], reduced male fertility, shortened hypocotyls, petioles and internodes, downward curled leaves, and delayed flowering
[82]. Moreover, BR-deficient plants also have a compact plant structure because of the reduced lamina inclination. However, BR deficiency can reduce grain size, seed fertility and tiller number
[9], improper stomatal distribution, and reduce seed germination
[86]. BR-insensitive and -deficient mutants are often referred to as late flowering due to their retarded growth
[87][88][87,88]. In contrast, plants over accumulating BRs display elongation of petioles and hypocotyls and increased height
[89].
Table 1. Involvement of various Brassinosteroid--regulated genes in plant growth and development.
|
Gene
|
Description of Gene
|
Crop/Plant
|
Role in Growth
|
Reference
|
|
CESA
|
The CESA gene superfamily, encoding the catalytic subunits of cellulose synthase
|
Arabidopsis
(A. thaliana)
|
Plays a role in regulating the cellulose synthesis
|
[90]
|
|
CYCD3;1
|
Cell division markers
|
Arabidopsis
(A. thaliana)
|
Needed for normal cell cycle progression
|
[51]
|
|
Histone lysine methyltransferase SDG8
|
In Arabidopsis, there are 43 SET Domain Groups (SDG), which contain proteins with conserved SET domains
|
Arabidopsis
(A. thaliana)
|
Involved in BR-regulated gene expression
|
[62]
|
|
WRKY46, WRKY54,
and WRKY70
|
The WRKY family TFs are composed of over 70 members in Arabidopsis
|
Arabidopsis
(A. thaliana)
|
Play positive roles in
BR-regulated plant growth and drought stress
|
[61]
|
|
Brassinazole-resistant 1 (BZR1), and BES1-interacting MYC-like proteins (BIMs)
|
BZR1; BR-activated transcription factor (TF) and BIMs; bHLH TF
|
Arabidopsis
(A. thaliana)
|
BR signaling promotes vegetative growth by inhibiting the floral transition
|
[91]
|
|
Transcripts of autophagy-related genes (ATGs)
|
Autophagy-related genes
|
Tomato
(Solanum lycopersicum)
|
Enhanced level of BR triggers ATGs and formation of autophagosomes
|
[92]
|
|
VvHMGR
|
Plays a role in the mevalonate (MVA) pathway
|
Grape berries
(Vitis vinifera)
|
Involved in increasing
the anthocyanin content and promoting coloration. Accumulates the fruit sugar components, and decreases the tartaric acid content
|
[93]
|