Porphyrias are a group of rare disorders originating from an enzyme dysfunction in the pathway of heme biosynthesis. Depending on the specific enzyme involved, porphyrias manifest under drastically different clinical pictures. The most dramatic presentation of the four congenital acute hepatic porphyrias (AHPs: acute intermittent porphyria—AIP, ALAD deficiency, hereditary coproporphyria—HCP, and porphyria variegata—VP) consists of potentially life-threatening neurovisceral attacks, for which givosiran, a novel and effective siRNA-based therapeutic, has recently been licensed. Nonetheless, the clinical manifestations of acute porphyrias are multifaceted and do not limit themselves to acute attacks. In particular, porphyria-associated kidney disease (PAKD) is a distinct, long-term degenerating condition with specific pathological and clinical features, for which a satisfactory treatment is not available yet. In PAKD, chronic tubule-interstitial damage has been most commonly reported, though other pathologic features (e.g., chronic fibrous intimal hyperplasia) are consistent findings. Given the relevant role of the kidney in porphyrin metabolism, the mechanisms possibly intervening in causing renal damage in AHPs are different: among others, δ-aminolevulinic acid (ALA)-induced oxidative damage on mitochondria, intracellular toxic aggregation of porphyrins in proximal tubular cells, and derangements in the delicate microcirculatory balances of the kidney might be implicated. The presence of a variant of the human peptide transporter 2 (PEPT2), with a greater affinity to its substrates (including ALA), might confer a greater susceptibility to kidney damage in patients with AHPs. Furthermore, a possible effect of givosiran in worsening kidney function has been observed. In sum, the diagnostic workup of AHPs should always include a baseline evaluation of renal function, and periodic monitoring of the progression of kidney disease in patients with AHPs is strongly recommended.
- porphyria
- kidney
- nephropathy
- chronic kidney disease
- kidney transplantation
- acute hepatic porphyrias
- porphyrin
1. Introduction
2. Role of the Kidney in Porphyrin Metabolism
3. Pathogenesis of Kidney Damage in PAKD
Among several mechanisms by which ALA is thought to cause cytotoxic damage, the kidney may be particularly susceptible—at least in its most metabolically active segments—to mitochondrial ALA-induced oxidation. At the intracellular level, ALA undergoes a phosphate-catalyzed auto-enolization, and becomes an oxidizing agent; it reacts with iron and O2 to produce superoxide anion (O2), HO radical, and ALA radical (ALA); ALA, in the presence of oxygen, reduces iron and yields dioxo valeric acid (DOVA), a highly reactive oxidant [26][27][36,37]. Several pieces of evidence have been gathered concerning ALA toxicity on mitochondrial morphology, loss of transmembrane potential, and protein expression [28][29][30][38,39,40]. Renal histopathological findings in patients with PAKD point toward chronic tubulointerstitial damage [16][18][31][32][33][34][16,18,30,41,42,43] and chronic fibrous intimal hyperplasia associated with focal cortical atrophy [16]. Early autopsy reports in a South African series of patients with variegate porphyria evidenced renal tubular degeneration, more marked in distal tubules, with calcified casts [18]. More recently, Pallet et al. [16] described tubular atrophy, basal membrane thickening, and interstitial fibrosis; nonspecific arteriosclerotic lesions [16] have also been observed, with arterial fibrous intimal hyperplasia in the cortex, consisting of myofibroblast growth, sclero-fibrotic tissue production and endothelial lumen narrowing. Remarkably, glomeruli seem spared from direct damage [34][43], since only unspecific sclerotic and ischemic lesions have been reported [16][31][16,30]. Markers of ongoing fibrogenesis, such as cytoplasmic accumulation of ß-catenin and vimentin expression, [16] have been detected in tubular sections, and mitochondrial abnormalities have been reported anecdotally [18][33][18,42]. Cell culture studies have shown that human endothelial cells (HUVECs), when incubated with ALA and PBG, do not appear to suffer direct damage from the porphyrin precursors [16]. In contrast, human renal epithelial cells (HRECs) display a wide range of alterations in the presence of ALA and PBG in vitro, i.e.: activation of apoptosis, with signs of autophagy and endoplasmic reticulum stress; evidence of a proinflammatory and fibrogenic secretory milieu; morphologic and molecular changes suggestive of epithelial-to-mesenchymal transition (loss of the cuboid morphology, cell-to-cell contact, E-cadherin expression; nuclear translocation of β-catenin; increased expression of SLUG). On electron microscopy, HRECs incubated with PBG showed accumulation of electron-dense cytosolic granules, whereas light microscopy detected yellow-brown granular aggregates, negative for Perl’s stain, and numerous cytoplasmic osmiophilic granules within the proximal tubular cells [16]. Intriguingly, when proximal tubular cells are incubated with PBG, the latter is completely metabolized into uroporphyrinogen I and III [16]: therefore, it has been conjectured that the observed intracellular inclusions could be aggregates of uroporphyrin obtained by the uncatalyzed polymerization and cyclisation of four PBG molecules. It is then interesting, from a historical as well as a scientific perspective, that a few studies on acute porphyrias from the past century have reported histopathological findings suggestive of tubular deposition of porphyrins [18][35][36][37][18,44,45,46]; for instance, a case series of autopsies from patients with variegate porphyria mentioned the presence of a brown autofluorescent pigment, not staining as iron, in both casts and renal tubular cells, and detected a red-orange autofluorescence in the lumen and epithelial cells of Henle’s loop, which in the author’s experience could be possibly attributed to porphyrin deposits [18]. As a matter of fact, consistent pieces of evidence have been gathered concerning the cell-damaging effects of light-independent porphyrin-mediated toxicity [38][47]: in particular, intracellular, extralysosomal porphyrin accumulation engenders protein aggregation through noncovalent, oxygen-dependent, reversible mechanisms [39][40][48,49]. A particular susceptibility has been demonstrated, chiefly in hepatocytes, for intermediate filaments (nuclear laminins and cytoplasmatic keratins) [38][41][47,50], proteins in the endoplasmic reticulum (e.g., protein disulfide isomerase and calnexin) [42][51], proteasome regulatory particles, and key glycolytic enzymes, including glyceraldehyde 3-phosphate dehydrogenase [42][51]. This process could both trigger and be accelerated by the activity of other oxidizing agents (inflammation, redox reactions) [39][48], so that porphyrins could precipitate the production of reactive oxygen species (ROS) and intracellular protein aggregation without prior photosensitization. Of note, uroporphyrin I is reduced by the P450 cytochrome’s family and by nicotinamide adenine dinucleotide phosphate (NADPH) in a reaction that yields a superoxide radical (O2−) [43][44][52,53]. It may be tempting to speculate that similar mechanisms might take place in the cytochrome-rich renal parenchyma, contributing to the renal toxicity of high concentrations of ALA and PBG. It must be remarked that when a mouse model of AIP was employed to investigate the effects of repeated phenobarbital-induced acute attacks on renal tissues [45][54] relatively mild unspecific alterations were undisclosed, even in near-total (that is, 5/6) nephrectomized animals. No granular inclusions or signs of tubule-interstitial damage were evidenced, even though the same authors underscore the differences between the experimental setting and the patients’ condition with years of exposure to abnormal levels of porphyrin precursors [45][54]. From a clinical perspective, signs of proximal tubulointerstitial insufficiency (i.e., proteinuria, impaired erythropoietin production) and of oxidative damage (increased urinary excretion of lipoperoxides), have been anecdotally signaled in porphyric patients [32][34][41,43]. A pattern consistent with sodium losses of tubular origin has been detected in patients with variegate porphyria [17]. A case series reported that, during remission from acute attacks, patients with AHPs displayed signs of tubulointerstitial and hypertensive damage, such as poorly concentrated urines (hyposthenuria), and an impairment of the tubular excretory phase, as disclosed by isotopic renography. In this population, four patients had low serum erythropoietin levels, while all of them (11 with AIP, 1 with VP) had low plasma and erythrocyte vitamin B6 (pyridoxal phosphate, PLP) levels. Interestingly, all patients had significant hyperoxalaemia and hyperoxaluria, and an inverse relationship between plasma oxalic acid and erythrocyte vitamin B6 levels was found in AIP patients [32][41]. Oxalic acid is a product of glyoxylic acid metabolism, whose conversion to glycine is effected by PLP-dependent transaminases [46][47][55,56]. Inherited excessive urinary excretion of oxalic acid (primary hyperoxaluria) is linked to an increased risk of urolithiasis (formation of calcium oxalate kidney stones) and kidney damage [48][57]. Even though the efficacy of PLP supplementation in reducing oxaluria is debated [47][48][49][50][51][56,57,58,59,60], AHPs patients are known to suffer from a poorer vitamin B6 status [52][53][61,62] compared to the general population4. Conclusions
The clinical manifestations of acute porphyrias are multifaceted and do not limit themselves to acute attacks. In particular, porphyria-associated kidney disease is a long-term, degenerating condition, for which a satisfactory treatment is still not available. A deeper understanding of the mechanisms of kidney damage in AHPs (Figure 1) is crucial for tailoring a treatment aimed at preventing progression to ESRD in these patients.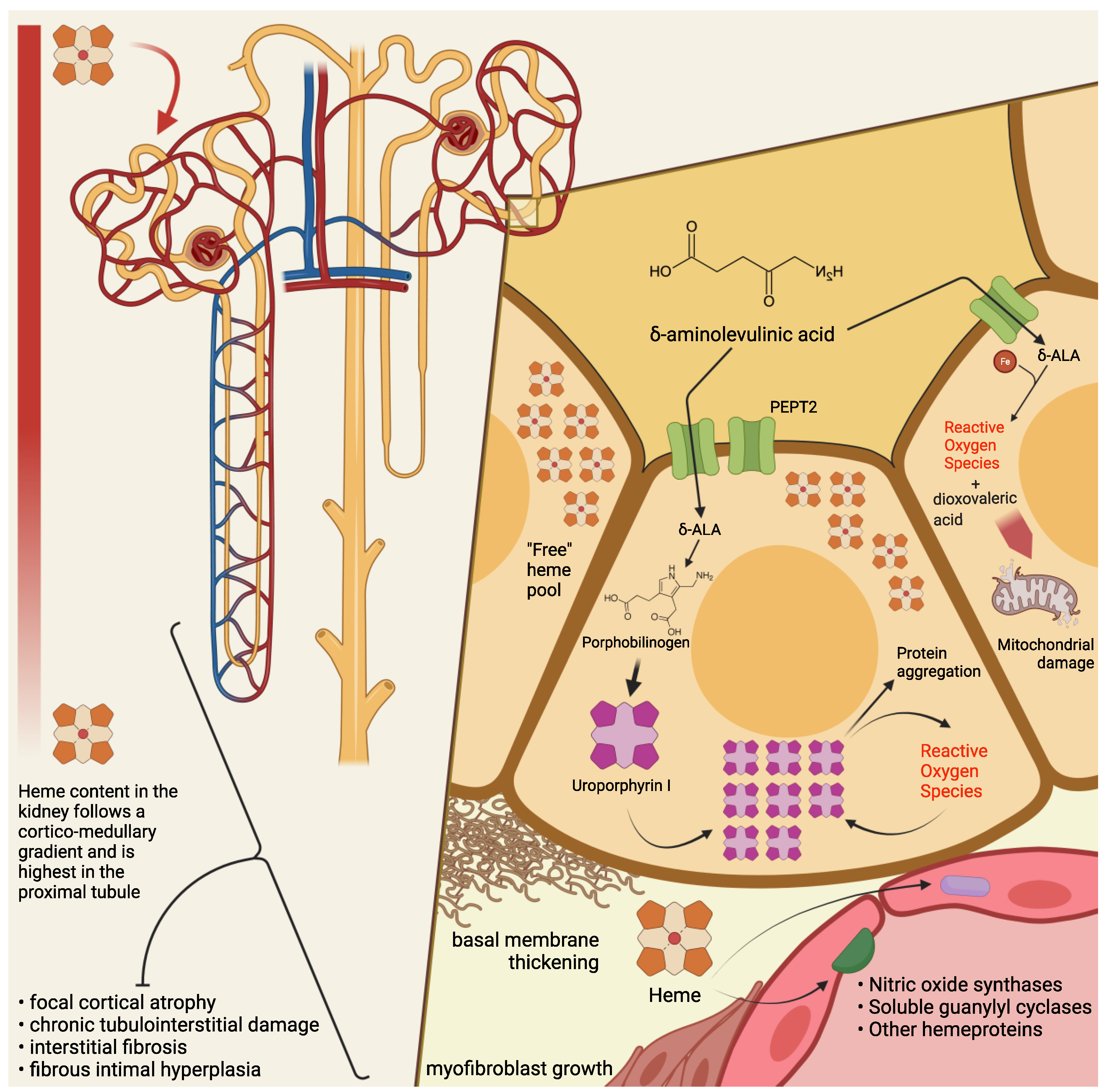
Acronyms
| AHP | Acute hepatic porphyrias |
| AIP | Acute intermittent porphyria |
| ALA | Aminolevulinic acid |
| ALAD | Aminolevulinic acid dehydratase |
| APA | Acute porphyric attack |
| CKD | Chronic kidney disease |
| DOVA | Dioxovaleric acid |
| eGFR | Estimated Glomerular Filtration Rate |
| ESRD | End-Stage Renal Disease |
| HCP | Hereditary coproporphyria |
| HMBS | Porphobilinogen-deaminase or hydroxymethylbilane-synthase |
| HREC | Human Renal Epithelial Cells |
| HUVEC | Human Umbilical Vein Endothelial Cells |
| mRNA | Messenger RNA |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NOS | Nitric oxide synthase |
| PAKD | Porphyria-associated kidney disease |
| PBG | Porphobilinogen |
| PEPT2 | Human Peptide Transporter 2 |
| PLP | Pyridoxal phosphate |
| RNA | Ribonucleic Acid |
| ROS | Reactive Oxygen Species |
| sGC | Soluble guanylyl cyclase |
| siRNA | Small interfering RNA |
| VP | Variegate porphyria |