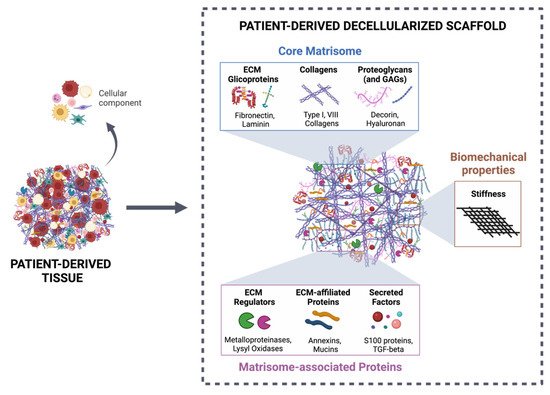
More than a physical structure providing support to tissues, the extracellular matrix (ECM) is a complex and dynamic network of macromolecules that modulates the behavior of both cancer cells and associated stromal cells of the tumor microenvironment (TME). Over the last few years, several efforts have been made to develop new models that accurately mimic the interconnections within the TME and specifically the biomechanical and biomolecular complexity of the tumor ECM. Particularly in colorectal cancer, the ECM is highly remodeled and disorganized and constitutes a key component that affects cancer hallmarks, such as cell differentiation, proliferation, angiogenesis, invasion and metastasis. Therefore, several scaffolds produced from natural and/or synthetic polymers and ceramics have been used in 3D biomimetic strategies for colorectal cancer research. Nevertheless, decellularized ECM from colorectal tumors is a unique model that offers the maintenance of native ECM architecture and molecular composition.
- colorectal cancer
- extracellular matrix
- patient-derived scaffolds
1. Introduction
2. Decellularized Colorectal Cancer Matrices as Bioactive Scaffolds for Modeling the Tumor Microenvironment
Decellularized ECM from malignant tissues is gaining attention in the field of organotypic modeling of tumor-stroma interactions by successfully incorporating key biochemical and biophysical characteristics of the native TME [27][28][29][133,134,135]. Particularly, patient-derived scaffolds allow comparisons between the tumor and the normal adjacent tissues, as well as deliver the potential of a preclinical platform to test patient-specific responses to treatment therapies [30][31][136,137]. However, decellularized ECM as a biomimetic model for CRC research is just beginning to be explored (Table 1) [24][25][24,25].| ECM Sources | Decellularization Method | Biochemical Evaluation | Biomechanical Evaluation | REF |
|---|---|---|---|---|
| Cell-derived matrix HT-29 SW480 CCD-841-Com |
-CHEMICAL 0.5% Triton X-100 20 mM NH4OH Ionic and nonionic surfactants |
n/a | n/a | [32][33][34][138,139,140] |
| Human-derived tissue | CHEMICAL 5 mM EDTA 10% DMSO 1% Triton X-100 10 mM sodium cholate hydrate 50 mM Tris-HCl Centrifugal rotation Ionic and nonionic surfactants Mechanical mixing |
-Cellular proteins (cytokeratin, vimentin) and stromal components (collagen IV, fibrinogen, hyaluronic acid): Immunohistochemistry -Actin: Western Blot -DNA content: SYBR agarose gel |
-Architecture: HE -3D structure: FITC staining of ECMs |
[35][50] |
| CHEMICAL/ENZYMATIC 4% sodium deoxycholate 2000 kU DNase-I |
-DNA content: DNeasy Blood & Tissue kit -Stromal components (GAGs, Col IV): PAS and Immunohistochemistry -Cellular proteins (Ki67, vimentin, E-cadherin, DAPI): Immunofluorescence |
-Architecture: HE and Laminin -3D structure: SEM -Permeability: In-house developed permeability device |
[36][49] | |
| -DNA content: DNeasy Blood & Tissue kit and 1% SYBRsafe agarose gel -Stromal components (GAGs, Col IV): PAS, Masson’s Trichrome, Immunohistochemistry and Alcian blue |
-Architecture: HE, Gieson and Silver stains -3D structure: SEM |
[19] | ||
| PHYSICAL/CHEMICAL Freezing 2% SDC 1% Triton X-100 Physical disruption Ionic and nonionic surfactants |
-Nucleic acids: HE -Collagens: SHG |
-Stiffness: AMR -Topography: SHG |
[37][52] | |
| CHEMICAL/ENZYMATIC 0.1% SDS 50 U/mL DNase-I Ionic surfactant |
-Nucleic acids: DAPI -DNA content: PureLink Genomic DNA Mini Kit -Histomorphological analysis: HE and Masson’s Trichrome -Major ECM proteins (Collagens I and IV, Laminin, Fibronectin and Hyaluronic acid): Immunofluorescence |
-Stiffness: Rheology -3D structure: SEM |
[26] | |
| CHEMICAL 1% SDS 1% Triton X-100 |
-DNA content: Nanodrop -Major ECM proteins (GAGs, Collagen I, Laminin and fibronectin): Immunostaining -Cellular proteins: F-actin (cytoskeleton), DAPI and HE (nuclei acid) |
-Structure and architecture: SEM and TEM | [38][141] | |
| SISmuc (small intestine submucosa + mucosa from decellularized porcine jejunum) |
CHEMICAL 4% SDS 200 U/mL DNase I- |
n/e | n/e | [39][142] |
| Mice-derived tissue | CHEMICAL/ENZYMATIC 4% sodium deoxycholate 2000 kU DNase-I |
-DNA content: Roche’s DNA isolation Kit and Quant-It PicoGreen dsDNA Assay -Nucleic acids: DAPI and HE -Major ECM proteins (Collagens I and IV, Fibronectin and Laminin): Immunofluorescence and Masson’s Trichrome |
-Tensile testing: RSA-G2 solids analyzer | [40][143] |