Obesity is caused by prolonged energy surplus. Current anti-obesity medications are mostly centralized around the energy input part of the energy balance equation by increasing satiety and reducing appetite. Our gastrointestinal tract is a key organ for regulation of food intake and supplies a tremendous number of circulating signals that modulate the activity of appetite-regulating areas of the brain by either direct interaction or through the vagus nerve. Intestinally derived messengers are manifold and include absorbed nutrients, microbial metabolites, gut hormones and other enterokines, collectively comprising a fine-tuned signalling system to the brain. After a meal, nutrients directly interact with appetite-inhibiting areas of the brain and induce satiety. However, overall feeding behaviour also depends on secretion of gut hormones produced by highly specialized and sensitive enteroendocrine cells. Moreover, circulating microbial metabolites and their interactions with enteroendocrine cells further contribute to the regulation of feeding patterns. Current therapies exploiting the appetite-regulating properties of the gut are based on chemically modified versions of the gut hormone, glucagon-like peptide-1 (GLP-1) or on inhibitors of the primary GLP-1 inactivating enzyme, dipeptidyl peptidase-4 (DPP-4). The effectiveness of these approaches shows that that the gut is a promising target for therapeutic interventions to achieve significant weigh loss.
- enteroendocrine cells
- enterokines
- gut microbiota
- pharmacological treatment
1. Introduction
Intestinally derived signals act on receptors on vagus nerves and neurons in appetite-controlling areas of the brain. In obesity, satiety signalling is inhibited, resulting in excessive food intake. Although a vast number of studies have been carried out to dissect the mechanisms that prevent the normal satiety in individuals living with obesity, the identity of the driving factor(s) is lacking. Most studies on obesity pathogenesis suggest the main cause is central dysregulation of feeding behaviour and lack of hunger suppression in the brain. Impaired gut hormone secretion and dysbiosis are often associated with obesity, but it is unclear whether this perturbed signalling has a causative role or whether its long-term perturbed signalling persists after weight loss and/or weight regain. Nevertheless, the signals derived from the intestine in the process of nutrient processing and uptake can be extremely powerful to supress the appetite. For example, the recent success of the GLP-1-based analogues for weight loss provides clear evidence of the potential for utilizing the intestinally-derived signalling pathway for obesity treatment. However, the intestine itself as a drug target can provide many more ways of modulating the energy intake.
Nutrient signalling is the primary signal initiating satiety at several levels and each one of them offers an opportunity to influence the satiety circuits—such as modulating the absorption rate or gut hormone secretion. Keeping in mind that the intestine is the largest inner organ in the body and contributes with 20–35% of whole-body energy expenditure [1], the diet-induced energy expenditure component is important for energy homeostasis. The intestinal epithelial lining is heavily vascularized and equipped with an underlying network of lymphatic capillaries which not only allow nutrient absorption but also interaction with immune cells. Recent studies show that these interactions are tightly connected to the satiety signalling. In addition, metabolites from symbiotic microorganisms, contributing to digestion, form signalling pathways to various systems of the human body, such as liver, immune system, and brain, helping to maintain energy homeostasis. As impaired regulation of food intake is the main driving force in obesity, focusing on origins of the natural satiety signalling and developing new ways of boosting it may provide us with better and safer ways of controlling the appetite.
2. Intestinal Epithelium and Signalling Properties of Nutrients
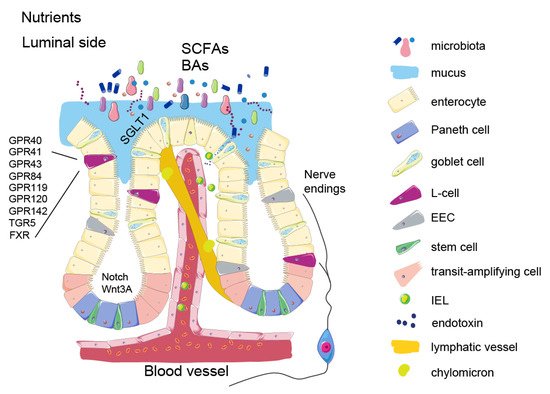
3. Enteroendocrine Cells
4. Microbial Metabolites
The microorganisms that colonize our gut, collectively referred to as the gut microbiota, are composed of roughly 39 trillion cells, comparable to the number of human cells in our body [17][78]. The combined genomes of the microbiota consist of more than 10 million genes, many of which encode enzymes that interact with the nutritional content of the gut to produce an extensive number of metabolically active compounds. These substances affect various host signalling pathways, including those controlling dietary intake and host metabolism. The majority of microbial species inhabit the distal gut, where, in the process of their livelihood, they produce metabolites beneficial for the host, such as vitamins, essential amino acids, short chain fatty acids (SCFAs), and bile acids (reviewed in Singh RK 2017). SCFAs, acetate propionate and butyrate, are a major energy source for colonocytes, but they can also act as signalling molecules. SCFAs can bind to GPR41 and GPR43 (FFAR3 and FFAR1) to stimulate secretion of GLP-1 from L-cells [18][19][79,80]. They also act as histone deacetylase (HDAC) inhibitors [20][81], key regulators of metabolism that increase thermogenesis in fat tissue. It has also been shown that propionate and butyrate can stimulate production and release of the anorexigenic hormone PYY from human EECs [21][82]. The effect of probiotic strains in obesity in human trials has shown some effects although the effect size generally has been moderate. Specifically, several studies using different strains of Lactobacillus, Streptococcus and Bifidobacterium have shown small effects on weight loss (<1 kg) [22][23][108,109], while others showed no difference in body weight after the treatment [24][110]. Furthermore, supplementation with pasteurized Akkermansia muciniphila enhanced weight loss more than live bacteria [25][106]. Although the described effects on weight loss seem rather subtle, these studies are encouraging as they demonstrated weight losses resulting from relatively short treatments and often by use of only a single, well-studied bacterial strain. As such, these studies can be viewed as encouraging proof-of-concept studies to inspire further studies with longer treatments, use of new bacterial strains and their combinations.5. Enterokines and Gut-Liver Axis
Non-endocrine enterocytes also produce factors with endocrine action, although these enterokines are not packed in large dense core granules, and their secretion is constitutive and do not require tightly regulated sensing machinery. Fibroblast growth factor (FGF) 15/19 (FGF19 in humans, FGF15 in rodents) is one of those enterokines. It is produced mostly in mature enterocytes in the ileum, and to a lesser extent in the jejunum and duodenum [26][27][111,112]. Many FGFs are growth factors with important functions for structural integrity of tissues, particularly of its mesenchymal component. However, some FGFs have low affinity for heparin sulphate glycosaminoglycans and thus are secreted into the circulation and as act as signalling molecules. These so called “endocrine” FGFs diffuse into the circulation and act on other organs in the body. This endocrine FGF signalling requires a co-receptor, with FGF15/19 specificity conferred via the FGFR1/beta klotho (KLB) receptor/co-receptor complex [28][113]. FGF15/19 production is induced when bile acids bind and activate the nuclear receptor FXR in the intestine. FGF15/19 functions as a signalling molecule by suppressing bile acid synthesis when bile acid levels are high in the intestinal mucosa, thus affecting bile receptor FXR signalling [29][30][31][32][114,115,116,117]. This bile acid-FGF15/19 signalling represses hepatic lipogenesis (which is abnormally high in obesity) [33][118] and inhibits gluconeogenesis in the liver [31][32][33][34][116,117,118,119]. When administered in supraphysiological levels or as pharmacological analogues, FGF15/19 also affects the adipose tissue and the brain, presumably through actions on FGFR1/KLB complexes [27][28][35][36][37][38][112,113,120,121,122,123].6. Intestinal Barrier Function
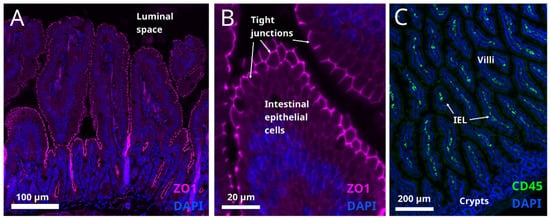
It is generally accepted that obesity is accompanied by low grade inflammation in the intestine [39][132]. The genesis of this condition is thought to be related to caloric overload. A study in humans shows that one month of high-fat feeding induces metabolic endotoxemia [40][133]. It is believed that high calorie foods sustain semi-pathogenic micro-organisms, and shift the balance away from the beneficial bacteria [41][134], mentioned above. Microbial pathogens can disrupt the tight junctions between the intestinal cells (Figure 34A) and thereby compromise the gut barrier function allowing the passage of endotoxins, which may induce systemic inflammation.