Cell-mediated immunity is driven by antigenic peptide presentation on major histocompatibility complex (MHC) molecules. Specialized proteasome complexes called immunoproteasomes process viral, bacterial, and tumor antigens for presentation on MHC class I molecules, which can induce CD8 T cells to mount effective immune responses. Immunoproteasomes are distinguished by three subunits that alter the catalytic activity of the proteasome and are inducible by inflammatory stimuli such as interferon-γ (IFN-γ). This inducible activity places them in central roles in cancer, autoimmunity, and inflammation.
- ubiquitin–proteasome system (UPS)
- immunoproteasome
- solid tumors
- proteasome inhibitors
1. Introduction
2. Structural and Functional Differences: Constitutive and Immunoproteasome
2.1. Composition, Assembly, and Regulation
3. Functional and Mechanistic Role of Immunoproteasome Subunits in Cancer
3.1. Role of Inducible Catalytic Subunits in Cancer
3.1. Role of Inducible Catalytic Subunits in Cancer
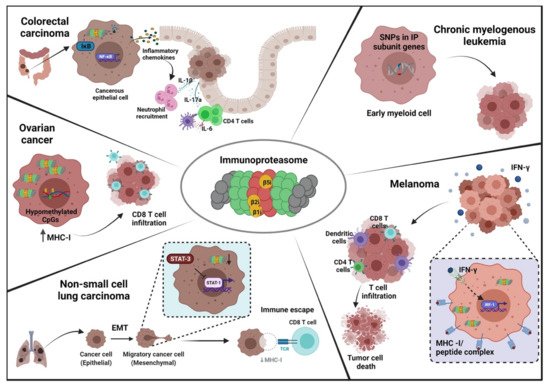
The generation of MHC class-I peptides is an important facet of the maturation of cytotoxic T cells (CTLs). Given the central function of CTLs in mounting anti-tumor responses, immunoproteasome subunits induced by IFN-γ have been studied for their assumed role in cancer development. Amongst the three IFN-γ inducible β subunits, β5i has to date been most implicated in blood and solid malignancies. This subunit, encoded by PSMB8 gene, has a wide range of expression among different cancers, which have been evaluated in non-small cell lung carcinoma (NSCLC), renal cell carcinoma, glioma, colorectal cancer, triple-negative breast carcinoma (TNBC), laryngeal, and hypopharyngeal carcinoma [21,22,148]. In many tumors, higher expression of PSMB8 has been linked with poor prognosis. PSMB8 expression was found to be upregulated in all histological sub-types of renal cell carcinoma [152]. Similarly, microarray profiling of gastric adenocarcinoma samples revealed that PSMB8 expression in tumor tissue was associated with poor prognosis [23]. High levels of PSMB8 are associated with more aggressive gliomas, and inhibition of PSMB8 was shown to reduce glioma cell proliferation and migration, as well to decrease glioblastoma tumor angiogenesis [148,153]. However, the observation that high expression of PSMB8 correlated with lower overall survival does not hold for all types of neoplasms. In NSCLC patients, high expression of PSMB8 was frequently observed in cancers with more favorable outcomes [22]. Likewise, increased PSMB8 expression in TNBC tumor samples was associated with better disease-free outcomes, including in those with metastatic disease [21].
The ambiguous role for PSMB8 in oncogenesis and disease progression seems to hinge on the fact that high levels of immunoproteasome expression can facilitate or impede tumor development in different contexts. For instance, the pro-tumorigenic role of PSMB8 in colorectal cancer is related to its role in colitis-induced chronic inflammation, which can drive neoplastic transformation of intestinal epithelium in the colon. Knockout of PSMB8 in mice was shown to prevent colitis-associated carcinogenesis [151]. PSMB8-deficient mice were found to be resistant to chronic inflammation and neoplasia, with reduced expression of chemokines CXCL-1, CXCL-2, and CXCL-3. Upon induction of colitis, PSMB8-/−
mice did not show macroscopic tumor development. The authors further attributed the pro-tumor effects of PSMB8 to reduced secretion of IL-17A in inflamed colons of PSMB8 deficient mice. The study proposed that IL-17A secretion was PSMB8-dependent via the NFκB signaling axis. The immunoproteasome has been shown to directly regulate NFκB signaling via direct proteolytic degradation of IκB, with knockdown of PSMB8 preventing the nuclear translocation of NFκB [85][86]. In its role in inflammation-driven carcinogenesis, PSMB8 serves as a promising treatment target for colorectal carcinomas. Supporting this, a study showed that ONX-914, an immunoproteasome inhibitor with a higher affinity for β5i subunit, suppressed tumor development in both preventive and therapeutic settings of colitis-induced carcinogenesis [87].mice did not show macroscopic tumor development. The authors further attributed the pro-tumor effects of PSMB8 to reduced secretion of IL-17A in inflamed colons of PSMB8 deficient mice. The study proposed that IL-17A secretion was PSMB8-dependent via the NFκB signaling axis. The immunoproteasome has been shown to directly regulate NFκB signaling via direct proteolytic degradation of IκB, with knockdown of PSMB8 preventing the nuclear translocation of NFκB [117,118]. In its role in inflammation-driven carcinogenesis, PSMB8 serves as a promising treatment target for colorectal carcinomas. Supporting this, a study showed that ONX-914, an immunoproteasome inhibitor with a higher affinity for β5i subunit, suppressed tumor development in both preventive and therapeutic settings of colitis-induced carcinogenesis [].
However, as mentioned, deficiency of PSMB8 is context-dependent. PSMB8 deficiency has been shown to promote tumor growth in a mouse model of melanoma. It has been observed that PSMB8-/−
mice implanted with B16 tumors have significant tumor growth and disease development [88]. In the absence of all three inducible subunits, mice failed to mount any anti-tumor immunity against the B16 melanoma cells, which was reflected in reduced CD8- mice implanted with B16 tumors have significant tumor growth and disease development [155]. In the absence of all three inducible subunits, mice failed to mount any anti-tumor immunity against the B16 melanoma cells, which was reflected in reduced CD8
+ T cells in the draining lymph nodes and CTLs in the tumor microenvironment (TME) and decreased IFN-γ expression [88]. This study postulated that in melanoma carcinogenesis, IFN-γ induced immunoproteasome expression by tumor cells increases infiltration of immune cells, further adding to the pool of cytokine and chemical mediators in the TME and further upregulating IFN-γ secretion, which can exert its anti-tumor functions. IFN-γ mediated overexpression of LMP7 in melanoma cells might increase the generation of neo-antigenic peptides, further accentuating an anti-tumor response. In support of this, overexpression of PSMB8 in melanoma cell lines increased IFN-γ secretion, leading to efficient killing of tumor cells by tumor infiltrating CTLs. This seemed to be mediated through the presentation of more diverse and immunogenic HLA-1 peptides generated through overexpression of immunoproteasome subunits [89].T cells in the draining lymph nodes and CTLs in the tumor microenvironment (TME) and decreased IFN-γ expression [155]. This study postulated that in melanoma carcinogenesis, IFN-γ induced immunoproteasome expression by tumor cells increases infiltration of immune cells, further adding to the pool of cytokine and chemical mediators in the TME and further upregulating IFN-γ secretion, which can exert its anti-tumor functions. IFN-γ mediated overexpression of LMP7 in melanoma cells might increase the generation of neo-antigenic peptides, further accentuating an anti-tumor response. In support of this, overexpression of PSMB8 in melanoma cell lines increased IFN-γ secretion, leading to efficient killing of tumor cells by tumor infiltrating CTLs. This seemed to be mediated through the presentation of more diverse and immunogenic HLA-1 peptides generated through overexpression of immunoproteasome subunits [156].
Thus, reduced expression of immunoproteasome subunits is a possible immune evasion mechanism deployed by tumor cells. In lung cancer, as in melanoma, higher expression of PSMB8 is associated with a more favorable prognosis, perhaps through increased immune surveillance [22,157]. In non-small cell lung carcinoma (NSCLC), tumor cells with lower expression of immunoproteasome subunits exhibited a more mesenchymal phenotype as opposed to the epithelial morphology of NSCLC cells with higher expression levels. Along with the mesenchymal phenotype, these tumor cells possessed increased migration and invasion ability with upregulated epithelial-to-mesenchymal transition (EMT) markers. Furthermore, STAT1 signaling was inhibited via the STAT3/mTOR regulatory axis in low PSMB8 expressing NSCLC cells. STAT1, in a mutually inhibitory relationship with STAT3, was shown to be a major downstream signaling molecule, controlling IFN-γ related genes including immunoproteasome and antigen presentation machinery. Upon treatment with IFN-γ, the mesenchymal phenotype of the tumor cells was reversed and phosphorylation of STAT1 was increased. Immunoproteasome induction in the mesenchymal-like NSCLC cell lines was shown to generate an increased diversity and quantity of MHC class I peptides. When pulsed with these generated peptides, autologous CD8 T cells demonstrated robust effector responses against tumor cells in vitro [157]. Thus, IFN-γ treatment induced immunoproteasome could potentially reverse this mechanism of tumoral immune evasion.
Immunoproteasomal subunit expression has also been reported to function as an indicator for treatment response and acquisition of chemoresistance. In both NSCLC and small cell lung cancer (SCLC), acquisition of cisplatin resistance correlated with increased expression of PSMB8 and PSMB9. Treatment of cisplatin-resistant tumor cells with proteasome inhibitors led to apoptosis induction, cell cycle arrest, and mitotic catastrophe. The authors propose that upregulation of immunoproteasome expression was a response to circumvent the cellular stress induced by cisplatin treatment [158]. Sensitivity to proteasome inhibitors by tumor cells was found to be associated with immunoproteasome subunit expression. In solid and hematological tumors, cells with low expression of immunoproteasome subunits showed poor response to proteasome inhibition, with significantly lower levels of apoptosis than cells with higher expression. However, pre-exposure with IFN-γ, which favored immunoproteasome subunit expression and immunoproteasome assembly, enhanced sensitivity to proteasome inhibitors [168]. As mentioned above, this raises a possibility of induction of the immunoproteasome through the IFN-γ pathway activation as a therapeutic strategy. For instance, resistance of the proteasome inhibitor bortezomib is associated with downregulation of PSMB8, which can be rescued through exogenous IFN-γ, leading to resensitization []. The resistance to inhibitor bortezomib was also found to be associated with mutation in the PSMB8 gene loci in multiple myeloma, which further potentiates the significance of screening PSMB8 mutations as well as expression for detecting chemoresistance to therapy [161].
A similar finding was noted in breast cancer. In TNBC, sensitivity to proteasome inhibitor treatment strongly correlated with high PSMB8 expression, with cells exhibiting UPS-driven apoptosis in response to immunoproteasome ablation [162]. To maintain high proliferative and invasive capacity, tumor cells increase protein turnover. Immunoproteasome upregulation by breast cancer cells is protective against increased proteotoxicity, which forms the part of unfolded protein response. Immunoproteasome upregulation, in this context, conceivably could be clinically targeted to overcome immunoproteasome driven chemoresistance, or could become a prognostic indicator of treatment responsiveness. Immunoproteasome expression was evaluated as a predictive marker for immune checkpoint blockade therapy in melanoma, with high expression of PSMB8 and PSMB9 associated with better response to anti-PD-1 and anti-CTLA-4 treatment [156].
PSMB8 expression in tumor cells not just reprograms the cellular pathways within the cell but also affects the tumor microenvironment. In highly invasive glioblastoma, a nexus of cellular communication is maintained between tumor cells, endothelial cells, and the extracellular matrix to allow increased angiogenesis. PSMB8 expression was reported to regulate this cellular communication. Elevated expression of PSMB8 was found in resected glioblastomas, and inhibition of PSMB8 reduced the migration and invasion of tumor cells in vitro. Endothelial cells demonstrated similar reduced migratory and tubulogenic properties when co-cultured with conditioned media taken from PSMB8-inhibited glioblastoma cell cultures. This interaction seemed to be mediated through reduced expression of vascular endothelial growth factor-A (VEGF-A) by tumor cells and integrin expression by endothelial cells. This was supported by a mouse model, which demonstrated that PSMB8 inhibition decreased tumor vessel formation [153]. However, the mechanism of VEGF-A control by the β5i subunit remains unexplained, with the authors hypothesizing immunoproteasome mediated degradation mechanism. Other studies have reported that PSMB8 regulation of migration and proliferation in less invasive grades of gliomas was dependent on PI3K and ERK pathways [149]. In addition to transcriptional and cytokine control of PSMB8 in cancer, regulation of PSMB8 has been reported through microRNAs, with miR-451a shown to target PSMB8 in prostate and thyroid cancer to prevent tumor cell proliferation and invasion [24,163].
The roles of the other two catalytic subunits, encoded by PSMB9 and PSMB10, are less described in cancer. β2i, or MECL1, encoded by PSMB10, has been reported to be downregulated in metastatic breast carcinoma, NSCLC, and acute promyelocytic leukemia however its functional relevance in tumor development is yet to be determined [157,164]. A recent study has implicated polymorphisms in PSMB10 as a genetic risk factor for chronic myelogenous leukemia (CML) []. The β1i subunit, encoded by PSMB9, was found to be reduced in breast cancer, renal cell carcinoma, APL, and NSCLC while elevated in melanoma and ovarian cancer [150,156,157,164,166–169]. Studies have reported a similar dichotomy as seen for PSMB8 regarding association with overall survival. Higher expression of PSMB9 in melanoma tumors has been linked with better patient outcomes while lower expression levels in NSCLC cells exhibited better prognosis [156,157]. A recent retrospective study on immune checkpoint therapy response for NSCLC and melanoma cohorts delineated a genetic signature of antigen processing and presentation (APM) genes which included PSMB9. Higher APM scores, and higher PSMB9 expression, correlated with better responses for immune checkpoint therapy (ICB) in both NSCLC and melanoma with improved overall survival [169].
The regulation of catalytic βi-subunits in cancer is brought about by several mechanisms. As described, NFκB, mTOR, and STAT1 have been shown to regulate the expression of PSMB8 in colon and lung cancer [154,157,]. In acute promyelocytic leukemia (APL), the fusion transcription factor PML/RARα resulting from the causative chromosomal rearrangement has been shown to interact with transcription factor PU.1 to repress the expression of all βi subunits [164]. As described above, contextual suppression of the immunoproteasome may provide a route for immune evasion, while upregulation may impart resistance to proteotoxicity. Aneuploidy, a common feature of neoplastic transformation, often increases protein production. Increased proteasomal degradation of tumor suppressor genes is another potential exacerbator of tumorigenesis. Constitutive proteasomal subunits are also frequently dysregulated during tumor initiation, and the induction of immunoproteasome subunits could provide extra capacity to cells undergoing intense protein turnover.
Apart from transcriptional control, immunoproteasome subunits are also regulated epigenetically. Hypomethylation of 6p21.3 CpG islands in high-grade serous epithelial ovarian carcinoma upregulates PSMB8/9 along with antigen presentation machinery proteins. This was found to be associated with increased time until recurrence time and increased CD8 T cell infiltration [167]. Low methylation profiles were observed for PSMB8 genomic regions in mucinous type epithelial ovarian cancers, which correlated with increased susceptibility to proteasome inhibitors [170]. Epigenetic modification of immunoproteasome subunits occurs diversely and is tumor-specific to which part of tumorigenesis it affects. Besides the regulation at the transcriptional and epigenetic level, immunoproteasome subunits themselves exhibit genetic polymorphisms which serve as susceptibility markers for certain cancers such as CML, cervical, and colon cancer [165, 171–172].
3.2. Role of Regulatory Subunits in Cancer
Even though immunoproteasome can process varied kinds of protein substrates, association with PA28 plays an important role in the generation of CTL-specific epitopes, by alerting conformation of the a rings [173–176]. While PA28 is also inducible by IFN-γ, it is also induced upon LPS or CD40 stimulation in dendritic cells [177]. The α and β subunits of the PA28 complex are differentially expressed and regulated independently. Since studies have shown that this dynamic expression influences clinical outcomes in various cancers, it has spiked the interest as to whether the differential expression and IFN-γ independent regulation of PA28 could independently promote the generation of tumor neoantigens.
In ovarian cancer, the C-terminal fragment of PA28 (PA28S or Reg-alpha, encoded by the PSME1 gene) was found in tumor biopsies with its presence correlated with poorer overall survival in patients, and was designated as a reliable biomarker to monitor tumor relapses and treatment [178]. Similarly, in multiple myeloma, the patients with a higher abundance of PA28α in their plasma showed reduced response to the proteasome inhibitor bortezomib [26]. Just as with the IFN-γ inducible catalytic subunits, the role of regulatory subunit expression in tumors is context-dependent. It has been reported that in oral squamous cell carcinoma (OSCC) and soft tissue leiomyosarcoma, high expression of PA28α in tumor samples corresponds with poor prognosis, while in melanoma, elevated levels of PSME1 were associated with better overall survival [25,27,179]. In OSCC cells, inhibition of PA28α in vitro led to decreased cell proliferation and a significant reduction in invasion ability and migration, implying a role in tumor growth and metastasis [179]. A similar role was shown in breast cancer cell lines, where PA28 inhibition was shown to increase CDK15, leading to suppression of migration and invasion [180]. Conversely, expression of PA28β was downregulated in esophageal squamous cell carcinoma, with overexpression inhibiting tumor cell proliferation in vitro [182]. However, there are limited functional studies to understand the mechanism underlying the differential behavior of PA28 and its subunits in cancer development. A recent study highlighted individual pathways of regulation for each subunit in cutaneous melanoma. Gene set enrichment and pathway-based analysis of the individual PSME genes showed independent and often contrasting pathways, for instance, PSME1 expression was positively correlated with increases in cell adhesion, apoptosis, and NFκB and Wnt signaling pathways while PSME2 was negatively correlated with the same [25]. PSME3 seemed to share features of PSME1 regulation, with correlation to NFκB and Wnt signaling pathways.
3.2. Role of Regulatory Subunits in Cancer
Besides its role as a prognostic marker, the PA28 complex also has been studied for its feasibility as a predictive marker for treatment response. PSME1 and PSME2 were included as part of the APM score described above that described responsiveness to ICB in NSCLC and melanoma [169]. This finding, however, is not consistent across all immunotherapies. PA28 was found to prevent effective responses in antigen-specific immunotherapy against melanoma. The protein MART-1 (also known as Melan- A or melanoma antigen recognized by T cells) has been investigated as a potential target for immunotherapy but initial trials showed a poorer than expected immune response. In vitro studies showed that the immunodominant MART-1 epitope was not efficiently recognized by CD8 T cells, due to epitope destruction by unexpected cleavage mediated by the PA28 complex [182]. In ICB, expression of the entire PA28 was observed to be a positive response marker. Alternatively, expression of PSME1 was found to be indicative of poor response to proteasome inhibitor treatment in relapsed or refractory multiple myeloma patients [26,169]. In another approach, PA28α was reported as an accessible target for therapeutic antibodies against prostate cancer [183].