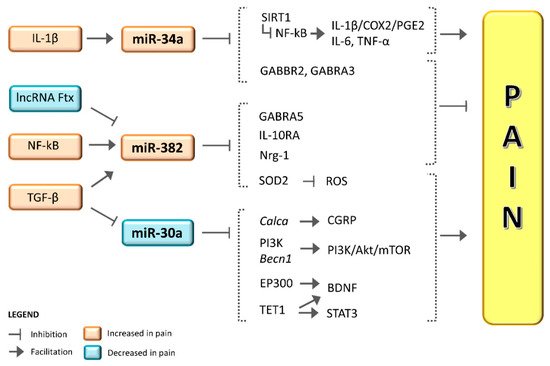
Preliminary but convergent findings suggest a role for microRNAs (miRNAs) in the generation and maintenance of chronic pain and migraine. Initial observations showed that serum levels of miR-382-5p and miR-34a-5p expression were increased in serum during the migraine attack, with miR-382-5p increasing in the interictal phase as well. By contrast, miR-30a-5p levels were lower in migraine patients compared to healthy controls. Of note, antimigraine treatments proved to be capable of influencing the expression of these miRNAs. Altogether, these observations suggest that miRNAs may represent migraine biomarkers, but several points are yet to be elucidated. A major concern is that these miRNAs are altered in a broad spectrum of painful and non-painful conditions, and thus it is not possible to consider them as truly “migraine-specific” biomarkers. We feel that Tthese miRNAs may represent useful tools to uncover and define different phenotypes across the migraine spectrum with different treatment susceptibilities and clinical features, although further studies are needed to confirm theour hypothesis. In this narrative entry , review we provide an update and a critical analysis of available data on miRNAs and migraines was provided, iin order to propose possible interpretations. TheOur main objective is to stimulate research in an area that holds promise when it comes to providing reliable biomarkers for theoretical and practical scientific advances.
- miRNA
- biomarker
- pain
- headache
- CGRP
1. Current evidence on miRNA expression in migraine.
| miRNA | Specimen | Population | Summary of Study Results | Reference | |||||
|---|---|---|---|---|---|---|---|---|---|
| Treatment | Posology and Duration | Population | miRNAs Modifications and Timing | Reference | |||||
| miR-382-5p | Serum | 8 migraine patients without any medication 12 migraine patients with normal medication habits 8 healthy controls |
4.1-fold increase expression during migraine attack. Higher expression during the interictal period in migraine patients vs. healthy subjects. |
Andersen et al. (2016) [1] | Andersen et al. (2016) [46] | ||||
| Detoxification | 7-day standardized detoxification protocol in hospitalized patients: abrupt withdrawal of overused drugs associated to intravenous therapy twice daily with isotonic 0.9% NaCl saline 500 mL + cyanocobalamin 2500 mcg + folic acid 0.70 mg + nicotinamide 12 mg + ascorbic acid 150 mg + sodic glutathione 600 mg + delorazepam 0.5 mg | 28 patients with CM–MOH | Significant reduction of miR-382-5p and miR-34a-5p two months after detoxification in CM–MOH. | Greco et al. (2020) [2] | Greco et al. (2020) [51] | ||||
| Peripheral blood mononuclear cells | 27 patients with EM 28 patients with CM–MOH |
Higher levels in CM–MOH vs. EM. Positive correlation with CGRP levels. |
|||||||
| Erenumab 70 mg | One administration s.c. every 28 days for a total of three administrations | 7 patients with CM 33 patients with CM–MOH | Greco et al. (2020) [2] | Greco et al. (2020) [51] | |||||
| Reduction of miR-382-5p and miR-34a-5p in the overall study population after three months of treatment with erenumab. | No differences between 30% Responders and NON-responders after three months of erenumab treatment. | De Icco et al. (2020) | [6] | De Icco et al. (2020) [52] | miR-34a-5p | Serum | |||
| NSAIDs and long-term magnesium | 8 migraine patients without any medication | Chronic treatment with magnesium (400 mg/day for three months) + Abortive treatment with acetaminophen (15 mg/kg) or ibuprofen (10 mg/kg) | 12 migraine patients with normal medication habits 8 healthy controls |
24 children and adolescents affected by migraine without aura divided into two groups (treated, and untreated) and 12 healthy controls | 9-fold increase expression during migraine attack. | Decreased expression of miR-34a-5p in migraine patients treated with NSAIDs and long-term magnesium vs. untreated migraine patients. Decreased expression in healthy controls vs. untreated migraine patients. Comparable expression between migraine patients treated with NSAIDs and long-term magnesium and healthy controls. | Andersen et al. (2016) [1] | Andersen et al. (2016) [46] | |
| Gallelli et al. (2019) | [ | 7 | ] | Gallelli et al. (2019) [50] | Peripheral blood mononuclear cells | 27 patients with EM and 28 with CM–MOH | Higher levels in CM–MOH vs. EM. Positive correlation with CGRP levels. |
Greco et al. (2020) [ | |
| Biphasic ketogenic diet | 2 | ] | Greco et al. (2020) [51] | ||||||
| Phase 1: 3 weeks with <30 g of carbohydrates | miR-30a | Serum | Patients with migraine with or without aura (sample size not defined) | Significant lower expression in patients with migraine vs. healthy controls. | Zhai et al. (2018) [3] | Zhai et al. (2018) [93] | |||
| Other miRNAs | Peripheral blood mononuclear cells | 15 female patients with migraine without aura 13 healthy controls |
Reduced miR-181a, let-7b and miR-22 levels in EM vs. HC. Increased miR-27b levels in EM vs. HC. |
Tafuri et al. (2015) [4] | Tafuri et al. (2015) [94] | ||||
| Serum | 8 migraine patients without any medication 12 migraine patients with normal medication habits 8 healthy controls |
Increased miR-29c-5p and miR-26b-3p expression in EM patients. | Andersen et al. (2016) [1] | Andersen et al. (2016) [46] | |||||
| Serum | 30 patients with EM 30 healthy controls |
Increased expression of miR-155, miR-126, and let-7 in EM vs. HC. | Chen et al. (2018) [5] | Chen et al. (2018) [95] |
2. miR-382-5p and miR-34a-5p in Migraine
3. miR-382-5p and miR-34a-5p in Pain and Putative Mechanisms