Fungi represent an essential component of ecosystems, functioning as decomposers and biotrophs, and they are one of the most diverse groups of Eukarya. In the tropics, many species are unknown. In this work, high-throughput DNA sequencing was used to discover the biodiversity of soil fungi in the Aguarongo forest reserve, one of the richest biodiversity hotspots in Ecuador. The rDNA metabarcoding analysis revealed the presence of seven phyla: Ascomycota, Basidiomycota, Mortierellomycota, Mucoromycota, Glomeromycota, Chytridiomycota, and Monoblepharomycota. A total of 440 identified species were recorded. They mainly belonged to Ascomycota (263) and Basidiomycota (127). In Mortierellomycota, 12 species were recorded, among which Podila verticillata is extremely frequent and represents the dominant species in the entire mycobiota of Aguarongo.
1. Introduction
The Andes represent the largest mountain range on Earth and cross the western part of South America. In Ecuador, the inter-Andean valley is composed of humid areas at high altitudes, with physiognomies ranging from grasslands to forest formations, combined with andisol soils, which develop from volcanic ash and show little or moderate evolution, forming a unique ecoregion. The forest soils in the Andean highlands have high biodiversity and unique environmental characteristics. It is an endangered ecosystem, and little is known about its microbial community. Fungi represent an essential functional component of terrestrial ecosystems as decomposers and biotrophs (mutualists, pathogens, and necrotrophs), and they are one of the most diverse groups of Eukarya
[1][2][1,2].
The soil fungal communities are affected by numerous biotic and abiotic factors, including seasons, soil characteristics, age and plant host species, and different soil managements
[3][4][3,4]. The study of the ecological factors that underlie the dynamics of fungal communities remains a challenge due to its high taxonomic and ecological diversity.
Molecular methods based on PCR and ribosomal DNA sequencing have been successfully used to identify fungi at different taxonomic levels in different environmental samples and have helped to elucidate the ecological conditions that affect the structure and diversity of fungal communities
[5][6][7][5,6,7]. The internal transcribed spacer region (ITS) is now widely used as a DNA barcode marker for the identification of fungal species
[8]. Recent studies have demonstrated the potential of advances in DNA sequencing for quantifying and characterizing the fungal diversity, especially in those areas of the planet where biodiversity is very rich and complex
[9].
Ecuador is one of the 17 megadiverse countries in the world, according to the World Conservation Monitoring Centre (WCMC) of the United Nations Environment Program. Ecuador has approximately 4.6 million hectares that have been declared as natural areas, which contain invaluable biodiversity and are a rich source of necessary ecological services. The fungal biodiversity of the natural areas in Ecuador is little known due to its extreme richness and complexity. The analysis of genetic material directly extracted from environmental samples coupled with DNA next-generation sequencing technology is useful in discovering its mycobiota and the monitoring of its biodiversity over time and in different ecological conditions
[10].
Rich literature can be found on the fungal biodiversity of Ecuador, mainly focused on taxonomy and distribution of a single taxon at the species or hierarchical higher level
[11][12][13][14][15][11,12,13,14,15]. Many papers focus on pathogenic fungi for important crops or on the relation between fungi and environments
[16][17][18][19][16,17,18,19]. Recently, environmental fungal DNA was analyzed for community composition studies of disturbed areas
[20][21][20,21] or specific substrates such as plants
[22][23][22,23]. Investigation of mycobiota of natural areas in Ecuador focused on biodiversity, distribution, and ecology of mycorrhizae
[24][25][26][24,25,26]. Data collected from environmental DNA concerning the entire mycobiota from natural areas and reserves of Ecuador are missing or rare.
2. Soil Chemical Characteristics
Concentrations of N, P, K, Mg, Ca, S, Cu, Mn, Zn, Fe, Na, exchangeable Al, and chlorides are presented in
Table 12 for each site. The analyses revealed that some chemical parameters vary significantly among the sampling sites; altitudinal levels could be one of the conditions that influence the chemical properties of the soil, but type of soil must be considered as well.
Table 12. Soil chemical properties. Values are expressed in mg/kg for each compound, except Al expressed in meq/100 g, while N and organic matter are expressed in % s.m.s. Values with different letters in the same column are significantly different (p < 0.05).
| Sampling Sites |
pH |
Organic Matter |
N |
P |
K |
Mg |
Ca |
S |
| L1 |
4.9 b |
10.9 a |
0.68 b |
3.6 b |
203.3 b |
425.7 b |
1776.3 b |
846.4 b |
| L2 |
4.8 a |
15.5 a |
0.82 a |
5.9 a |
128.0 a |
162.7 a |
564.7 a |
1012.6 a |
| L3 |
4.6 a |
12.3 a |
0.71 a |
5.3 a |
126.7 a |
177.7 a |
885.3 a |
790.4 a |
| Sampling Sites |
Cu |
Mn |
Zn |
Fe |
Na |
Cl |
Al |
|
| L1 |
10.5 b |
118.7 b |
16.0 a |
834.3 a |
28.7 a |
11.8 a |
6.4 a |
|
| L2 |
8.7 a |
72.7 a |
15.7 a |
926.0 a |
31.3 a |
15.1 a |
6.9 a |
|
| L3 |
12.7 a |
107.7 a |
16.0 a |
1197.3 b |
24.3 a |
371.8 b |
7.1 a |
|
The values of pH, N, P, K, Mg, Ca, S, Mn, and Cu are similar in L2 and L3 but are significantly different from L1. The content of Zn, Na, and exchangeable Al did not show differences between L1, L2, and L3. Site L1 showed the greatest differences, with lower values of N and P compared to the other locations, while in the case of Mg and Ca, it presents higher values.
3. Soil Fungal Assemblage Composition
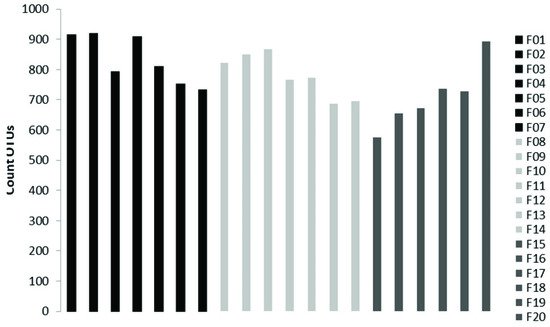
The sequencing on the Illumina MiSeq Platform of the DNA of the 20 pooled soil samples produced a total of 3,699,594 reads, with an average of 184,979.7 reads per sample. After the cutting and filtering process, the reads were reduced to 2,794,683, with an average of 139,734 reads per sample. It was observed that L1 showed the highest abundance with an average of 833 OTUs, followed by L2 with an average of 780 OTUs and L3 with an average of 711 OTUs (
Figure 12).
Figure 12. Number of OTUs present in each sample, with F1, F2, F3, F4, F5, F6, F7 belonging to altitudinal level L1, F8, F9, F10, F11, F12, F13, F14 beloning to L2, and F15, F16, F17, F18, F19, F20 belonging to L3.
Metagenomic data were taxonomically organized in this work following the high-level classification of the Fungi reported by Tedersoo et al.
[27][40]. The analysis revealed the presence of seven identified phyla in the soil samples:
Ascomycota and
Basidiomycota (subkingdom of
Dikarya),
Mortierellomycota,
Mucoromycota and
Glomeromycota (subkingdom
Mucoromyceta), and
Chytridiomycota and
Monoblepharomycota (subkingdom
Chytridiomyceta). The most abundant phylum was
Ascomycota, with a relative abundance of 32–36%, followed by
Mortierellomycota (25–28%),
Basidiomycota (9–11%), and
Mucoromycota (3–7%). The less abundant phyla were, as expected,
Glomeromycota, (0.1%)
Chytridiomycota (0.02%), and
Monoblepharomycota (0.0005%). The identified phyla were equally distributed in each sampling site, except
Glomeromycota, which was more abundant in Site L3, the highest in altitude. The share of fungal OTUs assigned to “Unidentified” was quite abundant in each site, reaching 17–35% of the total. In total, 408 taxa identified at the species level were found. All recorded species with their relative abundances in each sampling site are listed in
Supplementary Table S2 (available online at https://www.mdpi.com/article/10.3390/biology10121289/s1).
3.1. Subkingdom Dikarya
Ascomycota
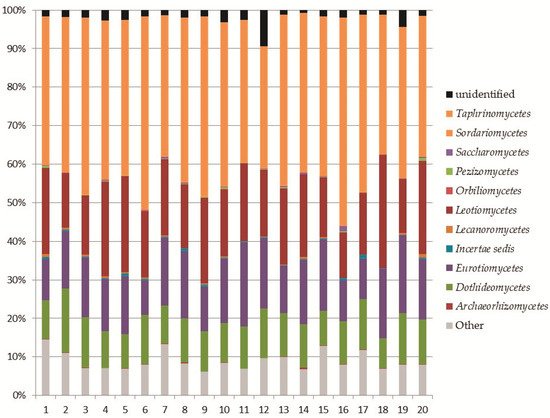
Within the phylum
Ascomycota, the analysis revealed the recording of 11 classes (
Figure 23). The most abundant classes were
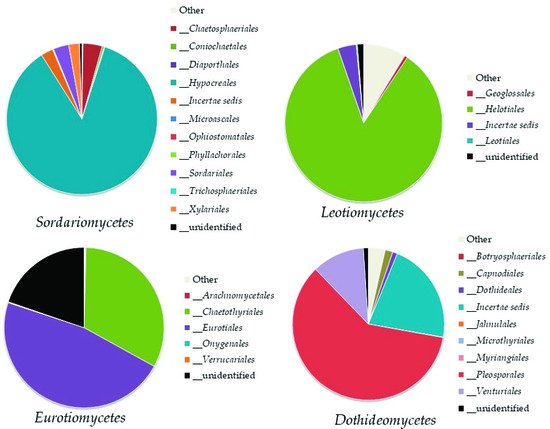
Sordariomycetes (8–17% of the total fungi), mainly represented by the orders
Hypocreales and
Chaetospheriales;
Leotiomycetes (3–9% of the total fungi), mainly represented by the order
Helotiales;
Eurotiomycetes (2–8% of the total fungi), mainly represented by
Eurotiales and
Chaetotyriales; and
Dothideomycetes (2–5% of the total fungi) mainly represented by
Pleosporales (
Figure 34).
Figure 23. Relative abundance of Ascomycota classes in the soil samples.
Figure 34. Order abundance in the most highly represented classes of Ascomycota: Sordariomycetes, Leotiomycetes, Eurotiomycetes, and Dothideomycetes.
In
Ascomycota, 263 identified genera were recorded; 58 genera reached at least 0.1% of the total fungi and were present in at least one of the samples with this value. Among them, the most abundant genera were:
Fusarium (6–12%),
Penicillium (0.5–6%),
Oidiodendron (0.1–2%),
Bipolaris (1–2%), and
Ilyonectria (0.4–1%). The other genera were recorded at less than 0.01%.
The ascomycetous taxa identified at the species level were 235 (
Supplementary Table S2). Most of them belonged to
Penicillium (nine species);
Trichoderma (six),
Oidiodendron (five),
Cladonia,
Cyphellophora,
Exophiala,
Peltigera, and
Phialophora (all with four species each), and
Acremonium, Aspergillus, Chalara, Chlonostachys, Hirsutella, and
Metarhizium (three species each).
Top Ten Most Abundant Species of Ascomycota
Among the identified taxa,
Fusarium oxysporum s. lat. was, undoubtedly, the most abundant species, reaching 7–10% of the total fungal assemblage.
Pleotrichocladium opacum was well represented (3%), together with
Penicillium spinulosum and
Curvularia lunata (2%). Other abundant identified species were
Trichoderma asperellum and
Ilyonectria panacis (1%),
Tetracladium apiense and
Chaetosphaeria vermicularioides (up to 0.5%),
Leptobacillium leptobactrum (up to 0.4%), and
Penicillium donkii (up to 0.3%).
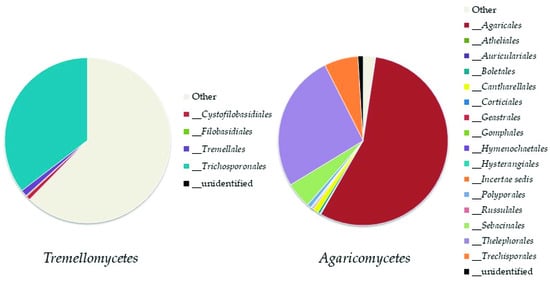
Basidiomycota
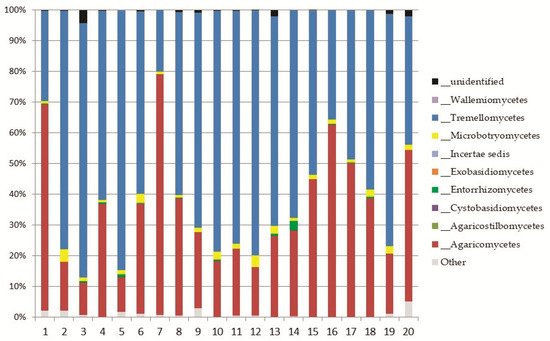
Basidiomycota was represented by eight classes (
Figure 45); the most abundant ones were
Tremellomycetes, represented by the order
Trichosporonales, and
Agaricomycetes (both, up to 11% of the total fungi), mainly represented by the orders
Agaricales and
Thelephorales (
Figure 56).
Figure 45. Relative abundance of classes of Basidiomycota in the soil samples.
Figure 56. Abundance of orders in the most represented classes of Basidiomycota: Tremellomycetes and Agaricomycetes.
The recorded genera of
Basidiomycota were 118. Only 29 of these genera reached at least 0.1% of abundance of the total fungi. Among them, the most abundant ones were:
Cuphophyllus,
Saitozyma, and
Trichosporon (up to 7% of the total fungi),
Tomentella (up to 6%), and
Hygrocybe (up to 2%).
The basidiomycetous taxa identified at the species level were 127 (
Supplementary Table S2). Most of the identified species belonged to
Entoloma (11 species), and
Lepiota (5 species).
Top Ten Abundant Species of Basidiomycota
The following species were the most abundant:
Saitozyma podzolica (3–4%),
Apiotrichum wieringae (1–2.5%), and Tomentella testaceogilva (1%).
Apiotrichum dulcitum,
Clitocybe nebularis,
Fomes fomentarius,
Porpolomopsis calyptriformis,
Sarcodon atroviridis, and
Serendipita vermifera were represented by less than 0.2%.
3.2. The Subkingdom Mucoromyceta
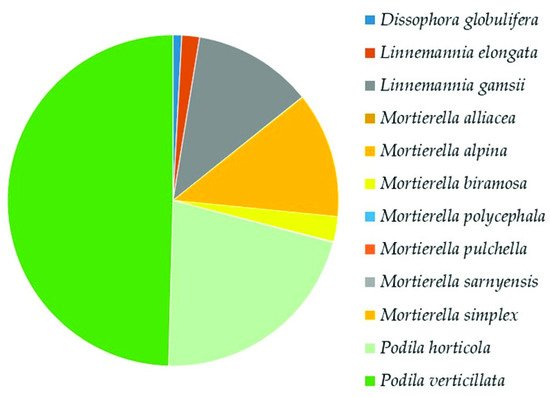
Mortierellomycota
Mortierellomycota was the second highest represented phylum. In the class
Mortierellomycetes, the analysis revealed the presence of 5 genera and 12 species (
Supplementary Table S2). The most dominant species were
Podia verticillata (50%),
Podia horticola (21%),
Mortierella alpina, and
Linnemannia gamsii (12% each) (
Figure 67).
Figure 67. Relative abundance of species in
Mortierellomycetes (
Mortierellomycota), following the most recent taxonomic classification at the species level
[28][41].
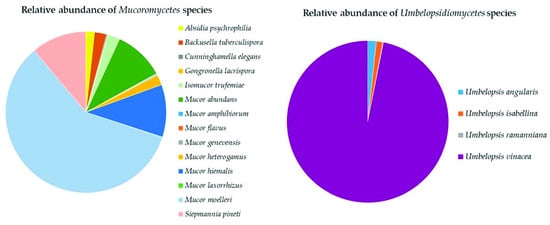
Mucoromycota
The three classes of
Mucoromycota were all represented in the soil samples with a total of 19 species (
Supplementary Table S2).
Mucoromycetes (14 species) were dominated by
Mucor moelleri (58%),
Mucor hiemalis,
Sepmannia pineti (both 11%), and
Mucor abundans (10%) (
Figure 78).
Umbelopsidiomycetes (four species) was completely dominated by
Umbelopsis vinacea (97%,
Figure 78).
Endogonomycetes was present with only one identified species,
Jimgerdemannia lactiflua.
Figure 78. Relative abundance of species of Mucoromycota in the classes Mucoromycetes and Umbelopsidiomycetes.
Glomeromycota
The taxa belonging to this phylum were poorly identified at the species level. The detected species were seven (
Supplementary Table S2), belonging to six genera:
Claroideoglomus claroideum (2%) followed by
Acaulospora alpina, A. lacunosa, Racocetra castanea, R. fulgida, Archaeospora trappei, and
Dentiscutata heterogama.
The Top Mucoromyceta
Of the total fungi,
Podila verticillata was strongly represented (12–14%), followed by
P. horticola (5–6%),
Mortierella alpina (2–6%),
Linnemannia gamsii (3%),
Umbelopsis vinacea (2.5–3%),
Mortierella biramosa (0.5–1%), and
Mucor moelleri (0.1–1%).
3.3. The Subkingdom Chytridiomyceta
In
Chytridiomyceta, the identified species were very rare (four, listed in
Supplementary Table S2). The most abundant was
Rhizophlyctis rosea (13%), belonging to
Chytridiomycota, followed by
Spizellomyces plurigibbosus (6%),
Olpidium brassicae (1%), and
Gallinipes pseudodichotomus (1%). In
Monoblepharomycota, the only identified species was
Monoblepharella mexicana (3%).
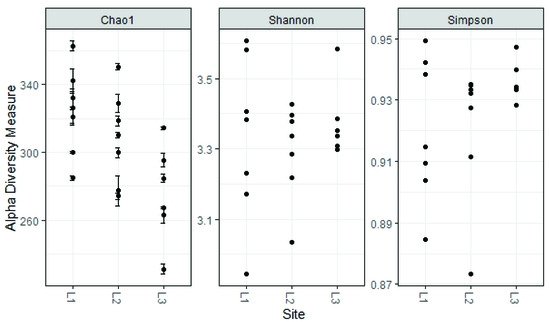
4. Soil Mycobiota Diversity in the Sampling Sites
The alpha diversity found in the fungal communities of the three locations (L1, L2, L3) was similar regarding the Shannon and Simpson Indices, while the Chao 1 Index, which focuses especially on the abundance of rare species, found a significant difference between locations L1 and L3 (324 ± 26 and 275 ± 29, respectively,
p < 0.05 ANOVA, followed by Tukey’s pairwise test,
Figure 89).
Figure 89. Estimated alpha diversity for different sampling sites.
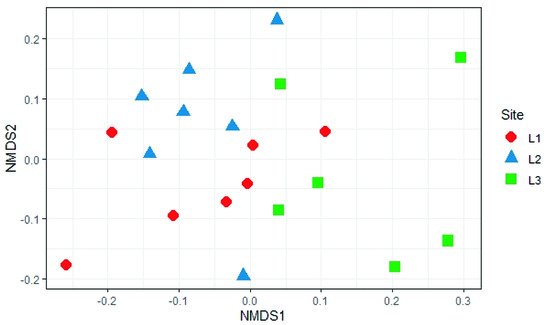
The analysis of beta diversity present in the samples did not show any significant difference in the structure of soil fungal communities in the three sampling locations (PERMANOVA,
p > 0.05;
Figure 910).
Figure 910. Non-Metric Multidimensional Scaling (NMDS) based on Bray-Curtis distances of soil fungal communities in the different sample locations (L1, L2, and L3) in the Aguarongo forest.
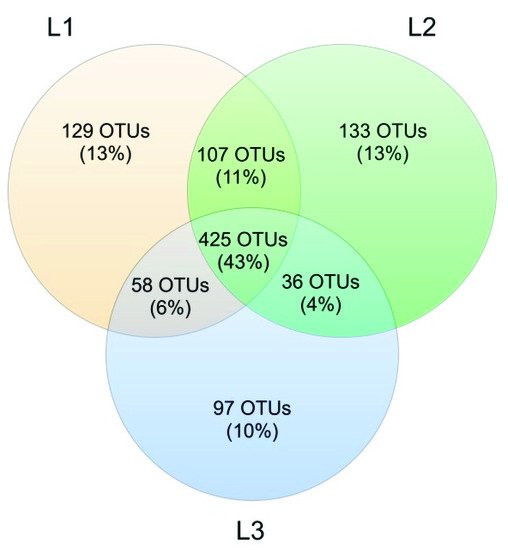
Venn’s analysis showed that 43% of all OTUs was shared between the three sampling sites. At Locations L1 and L2, the number of unique OTUs was similar (129 and 133, respectively), while L3 had a lower number of unique OTUs (97, corresponding to 10%) (
Figure 101).
Figure 101. Venn diagram showing OTUs shared among the 3 sampling sites (L1, L2, and L3).
5. Discussion
Discussion
The Aguarongo forest reserve is one of the most important biodiversity hotspots in Ecuador, and in recent years, it has been the subject of numerous research studies on flora, fauna, use of soils, and water balance [29][30].The Aguarongo forest reserve is one of the most important biodiversity hotspots in Ecuador, and in recent years, it has been the subject of numerous research studies on flora, fauna, use of soils, and water balance [28, 42].
As for all the tropics, Ecuador has huge biodiversity that has yet to be revealed, especially in natural reserves such as the Aguarongo forest. This is true above all for Fungi, a Kingdom often neglected despite its key role in the functionality of terrestrial ecosystems.
Regarding the use of next-generation sequencing in research of soil fungal communities, there are few studies in the tropics; however, a collection of metagenomic data is being implemented [31][32][33][34]. Most Ecuadorian studies on soil fungi describe the diversity of arbuscular mycorrhizal fungi (AMF) associated with crops in the Andean part of Loja and Quito [35][36]. There are also studies of the communities of AMF associated with plants growing in contaminated soils such as crude-oil-contaminated sites in the Amazon region of Ecuador [20]. Using modern molecular methods, it was possible to obtain a detailed account of soil fungal biodiversity of the Aguarongo forest and give the first list of soil fungi of this important reserve of biodiversity in Ecuador.Regarding the use of next-generation sequencing in research of soil fungal communities, there are few studies in the tropics; however, a collection of metagenomic data is being implemented [43–46]. Most Ecuadorian studies on soil fungi describe the diversity of arbuscular mycorrhizal fungi (AMF) associated with crops in the Andean part of Loja and Quito [47,48]. There are also studies of the communities of AMF associated with plants growing in contaminated soils such as crude-oil-contaminated sites in the Amazon region of Ecuador [20]. Using modern molecular methods, it was possible to obtain a detailed account of soil fungal biodiversity of the Aguarongo forest and give the first list of soil fungi of this important reserve of biodiversity in Ecuador.
In this work, 408 fungal OTUs were identified at the species level. Together with Ascomycota, the most abundant phylum in our study was Mortierellomycota. Its high abundance was expected, as this result has been recorded in other studies on the mycobiota of soil in high altitude environments [32][34] or from extreme environments such as the Alps [37] or Antarctica [38][39][40]. Surprisingly, in the Aguarongo forest, the abundance of Mortierellomycota is similar to that of Ascomycota. All the OTUs referring to Mucoromyceta in this study were updated following the recent paper of Vandepol et al. [28], in which the phylogeny of Mortierellaceae was resolved through a synthesis of multigene phylogenetics and phylogenomics. The present work significantly improved the global geography and environmental records of Mortierellaceae and contributed to the knowledge on the ecology of these species, especially in an under-sampled region such as Ecuador. These data contribute to understanding the ecological function of Mortierellaceae, which remains mostly unknown. Species of Mortierellaceae are often isolated from soils, decaying leaves, and insects [41], due to their saprotrophic nature [42].In this work, 408 fungal OTUs were identified at the species level. Together with Ascomycota, the most abundant phylum in our study was Mortierellomycota. Its high abundance was expected, as this result has been recorded in other studies on the mycobiota of soil in high altitude environments [44,46] or from extreme environments such as the Alps [49] or Antarctica [50–52]. Surprisingly, in the Aguarongo forest, the abundance of Mortierellomycota is similar to that of Ascomycota. All the OTUs referring to Mucoromyceta in this study were updated following the recent paper of Vandepol et al. [41], in which the phylogeny of Mortierellaceae was resolved through a synthesis of multigene phylogenetics and phylogenomics. The present work significantly improved the global geography and environmental records of Mortierellaceae and contributed to the knowledge on the ecology of these species, especially in an under-sampled region such as Ecuador. These data contribute to understanding the ecological function of Mortierellaceae, which remains mostly unknown. Species of Mortierellaceae are often isolated from soils, decaying leaves, and insects [53], due to their saprotrophic nature [54].
Regarding alpha diversity, it was similar among all sampling sites; the values reported here are similar to those found for other regions of the world with altitudes above 2000 m.a.s.l. [32].Regarding alpha diversity, it was similar among all sampling sites; the values reported here are similar to those found for other regions of the world with altitudes above 2000 m.a.s.l. [44].
In the whole soil mycobiota of Aguarongo, the most frequent species is a Mortierellaceae species, Podila verticillate, representing 12–14% of the total of identified OTUs. This species (reported with its basionym Mortierella verticillata) was first recorded in South America from mountainous environments (1950 m.a.s.l.) in Brazil [43]. Our findings expand the distribution of P. verticillata both in latitude and altitude. This is true for all of the species of Mortierellaceae, which were abundantly found in Aguarongo forest soil up to 3221 m.a.s.l.In the whole soil mycobiota of Aguarongo, the most frequent species is a Mortierellaceae species, Podila verticillate, representing 12–14% of the total of identified OTUs. This species (reported with its basionym Mortierella verticillata) was first recorded in South America from mountainous environments (1950 m.a.s.l.) in Brazil [55]. Our findings expand the distribution of P. verticillata both in latitude and altitude. This is true for all of the species of Mortierellaceae, which were abundantly found in Aguarongo forest soil up to 3221 m.a.s.l.
Within the identified species of Ascomycetes in Aguarongo, important facultative plant pathogenic fungi were abundantly recorded, for example from the Fusarium oxysporum complex, a well-known group of taxa due to their many pathogenic forms and high frequency of isolation in soil [44], and Curvularia lunata, an important pathogen of maize [45]. In addition to the plant pathogens many saprotrophic fungi were recorded, such as Pleotrichocladium opacum and Penicillium spinulosum, a dominant taxon in the Aguarongo forest.Within the identified species of Ascomycetes in Aguarongo, important facultative plant pathogenic fungi were abundantly recorded, for example from the Fusarium oxysporum complex, a well-known group of taxa due to their many pathogenic forms and high frequency of isolation in soil [56], and Curvularia lunata, an important pathogen of maize [57]. In addition to the plant pathogens many saprotrophic fungi were recorded, such as Pleotrichocladium opacum and Penicillium spinulosum, a dominant taxon in the Aguarongo forest.
Beneficial fungi were not lacking, such as Trichoderma asperellum, an important biocontrol agent, applied in agriculture for controlling plant fungal pathogens and plant-growth promoters [46][47], as well as Arthrobotrys musiformis Pochonia bulbillosa, and P. suchlasporia, well-known nematophagous fungi [48][49]. Particularly important in the Aguarongo forest is the presence of the genus Metarhizium, a genus that shows wide biodiversity in this area, with species having high potential as biocontrol agents against insects, specifically M. anisopliae, M. flavoviride, and M. lepidiotae, [50]. Although their presence is occasional, many other species with importance because of their ecological role as entomopathogens or nematopathogens, and potential application for sustainable agriculture [51] are found in the Aguarongo forest. This includes Beauveria brongniartii, B. bassiana, Dactylella mammillata, Hirsutella minnesotensis, H. rhossiliensis, H. vermicola, Paecilomyces farinosus, and P. marquandii.Beneficial fungi were not lacking, such as Trichoderma asperellum, an important biocontrol agent, applied in agriculture for controlling plant fungal pathogens and plant-growth promoters [58,59], as well as Arthrobotrys musiformis Pochonia bulbillosa, and P. suchlasporia, well-known nematophagous fungi [60,61]. Particularly important in the Aguarongo forest is the presence of the genus Metarhizium, a genus that shows wide biodiversity in this area, with species having high potential as biocontrol agents against insects, specifically M. anisopliae, M. flavoviride, and M. lepidiotae, [62]. Although their presence is occasional, many other species with importance because of their ecological role as entomopathogens or nematopathogens, and potential application for sustainable agriculture [63] are found in the Aguarongo forest. This includes Beauveria brongniartii, B. bassiana, Dactylella mammillata, Hirsutella minnesotensis, H. rhossiliensis, H. vermicola, Paecilomyces farinosus, and P. marquandii.
Basidiomycetes were dominated by Saitozyma podzolica, a common yeast and strong decomposer of dead plant biomass isolated from soils worldwide [52]. Apiotrichum wieringae was also widely present in the soil of Aguarongo, a nonpathogenic member of the Tricosporonaceae family, able to degrade uric acid and aromatic compounds [53]. Tomentella testaceogilva, abundant in Aguarongo, is reported as a terricolous or lignicolous fungus associated with Alnus, but also with moss [54]. Although rarely recorded in the Aguarongo forest, a famous member of Basidiomycetes must be highlighted, namely Fomes fomentarius, an important plant pathogenic fungus rich in pharmacological compounds [55].Basidiomycetes were dominated by Saitozyma podzolica, a common yeast and strong decomposer of dead plant biomass isolated from soils worldwide [64]. Apiotrichum wieringae was also widely present in the soil of Aguarongo, a nonpathogenic member of the Tricosporonaceae family, able to degrade uric acid and aromatic compounds [65]. Tomentella testaceogilva, abundant in Aguarongo, is reported as a terricolous or lignicolous fungus associated with Alnus, but also with moss [66]. Although rarely recorded in the Aguarongo forest, a famous member of Basidiomycetes must be highlighted, namely Fomes fomentarius, an important plant pathogenic fungus rich in pharmacological compounds [67].
Fungi in soils of Aguarongo include members of the phylum Glomeromycota, which remain mainly unidentified. Some identified species are important arbuscular mycorrhizal fungi; the most abundant one was Claroideoglomus claroideum, considered a good plant promoter and heavy metal decontaminant [56]. In the Aguarongo forest, Chytridiomycota were dominated by Rhizophlyctis rosea, a highly effective plant biomass degrader and zoosporic fungus, commonly observed near the soil surface. This species has light-sensitive proteins that allow this fungus to remain in the euphotic zone [57]. Within Monoblepharomycota, only one species was recorded: Monoblepharella mexicana. The southernmost record of this species was reported by Steciow and Arambarri [58], while the present finding can be considered the record of this species at the highest elevation.Fungi in soils of Aguarongo include members of the phylum Glomeromycota, which remain mainly unidentified. Some identified species are important arbuscular mycorrhizal fungi; the most abundant one was Claroideoglomus claroideum, considered a good plant promoter and heavy metal decontaminant [68]. In the Aguarongo forest, Chytridiomycota were dominated by Rhizophlyctis rosea, a highly effective plant biomass degrader and zoosporic fungus, commonly observed near the soil surface. This species has light-sensitive proteins that allow this fungus to remain in the euphotic zone [69]. Within Monoblepharomycota, only one species was recorded: Monoblepharella mexicana. The southernmost record of this species was reported by Steciow and Arambarri [70], while the present finding can be considered the record of this species at the highest elevation.
The analysis reported in this work is based on a large number of soil samples (100), thus giving a good representation of the fungal biodiversity of Aguarongo forest soil. The collection of samples along an altitudinal gradient did not reveal significant differences among the structure of the soil fungal communities in the three sampling locations (L1, L2, L3). Only Location L3 has a lower abundance of rare taxa with respect to L1, consistent with the Venn diagram.
The analysis of the records, reported in Supplementary Table S2, shows the Aguarongo forest soils as a natural area rich in fungal biodiversity, with the main phyla represented by a large number of taxa, known to be beneficial fungi. In addition, the Aguarongo forest is shown to be a rich source of species having strong application potential in agriculture and in the chemical and food industry. This area hides a huge number of unknown species that could be assessed, and its protection is of the utmost importance.
The analysis of the records, reported in Supplementary Table 2, shows the Aguarongo forest soils as a natural area rich in fungal biodiversity, with the main phyla represented by a large number of taxa, known to be beneficial fungi. In addition, the Aguarongo forest is shown to be a rich source of species having strong application potential in agriculture and in the chemical and food industry. This area hides a huge number of unknown species that could be assessed, and its protection is of the utmost importance.