Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Vivi Li and Version 1 by Carla Maleita.
Rice (Oryza sativa L.) is one of the main cultivated crops worldwide and represents a staple food for more than half of the world population. Root-knot nematodes (RKNs), Meloidogyne spp., and particularly M. graminicola, are serious pests of rice, being, probably, the most economically important plant-parasitic nematode in this crop. M. graminicola is an obligate sedentary endoparasite adapted to flooded conditions. Until recently, M. graminicola was present mainly in irrigated rice fields in Asia, parts of the Americas, and South Africa. However, in July 2016, it was found in Europe, in northern Italy in the Piedmont region and in May 2018 in the Lombardy region in the province of Pavia.
- damage
- hosts
- life cycle
- plant-parasitic nematode
- rice root-knot nematode
1. Introduction
Rice (Oryza sativa L.) is the third most important cereal crop in the world, just behind wheat and maize, playing a strategic role in solving food security issues. New risks to plant health are constantly emerging. Many nematodes in rice have been detected and described, but only a few have harmful effects on rice production, such is the case of the rice root-knot nematode (RKN) Meloidogyne graminicola Golden and Birchfield, 1965 (Mg) [1], recently detected in Italy and added to the European and Mediterranean Plant Protection Organization (EPPO) Alert List [2]. Mg is considered a major threat to rice production, particularly in Asia. Projections by the Intergovernmental Panel for Climate Change indicate that there will be an increase in mean annual temperature and rainfall in South Asia, West Africa, and Europe. The elevated temperature and moisture may result in an increasing rate of infection, development, and reproduction, causing shifts in Mg abundance and geographic distribution. Such effects may have a detrimental impact on rice in temperate regions. Furthermore, Mg is a clear example of how alterations in rice production (shortage of water due to socioeconomic pressure and climate change) contributed to changes in its status as the major plant-parasitic nematode (PPN) in rice. An effort has been made to gather all the information regarding several aspects of Mg to present it as a comprehensive review on rice RKN.
2. Origin and Distribution
The rice RKN, Mg, was first isolated in India by Israel et al. [3], but it was only described in 1965 when it was found on the roots of barnyard grass (Echinochloa colonum) in Baton Rouge, Louisiana, USA [4]. Since then, this nematode has been reported from the USA on rice and weeds in Louisiana, on grass in Georgia and Mississippi, and on sandbur (Cenchrus spp.) in Florida [5,6,7,8][5][6][7][8]. Its occurrence has been widely accounted in rice fields in several Asian countries [9,10,11][9][10][11] and also in South Africa, Colombia, Brazil, and Italy [12,13,14][12][13][14]. Mg has been reported to parasitize primarily in irrigated and rainfed rice in South and Southeast Asian countries, such as China, India, the Philippines, Burma (Myanmar), Bangladesh, Pakistan, Laos, Thailand, Vietnam, and Nepal [15,16,17][15][16][17]. In China, it was first found on Allium tistulosum in the Hainan province by Zhao et al. [18]. More than a decade later, it was detected associated with rice and other hosts including weeds in the provinces of Anhui, Fujian, Hainan, Hunan, Hubei, Zhejiang, Jiangxi, and Sichuan, causing a severe incidence in the Hunan province [19,20,21,22][19][20][21][22]. In India, this nematode was first isolated in the county of Orissa from upland rice soils by Israel et al. [3]. Since then, it has been found infecting rice in the provinces of Andaman and Nicobar Islands, Assam, Andhra Pradesh, Bihar, Gujarat, Himachal Pradesh, Jammu and Kashmi, Karnataka, Kerala, Madhya Pradesh, Manipur, Orissa, Tamil Nadu, Tripura, and West Bengal [23,24][23][24]. In 1971, its presence was referred in Thailand, causing typical root galls in entire rice-growing areas and in nursery seedbeds [25], and in Bangladesh, where it has been often associated with deepwater and pre-monsoon upland rice systems [26,27,28][26][27][28]. Minor infestations were reported in lowland rainfed rice areas [28]. Nonetheless, in the northwest of Bangladesh, where the dominant cropping system is lowland rainfed alternated with wheat, severe infestations of Mg were observed [29]. Later, in the 1990s, Mg was reported infesting rice fields in Sri Lanka, where it is now dispersed into major rice-growing areas of the country [30,31,32][30][31][32]. In a study performed in Vietnam, in 1992, to determine the PPN in deepwater rice systems, Mg was identified for the first time as one of the main causes of high yield losses of rice [33]. In Pakistan, during a survey in rice fields of Sheikhupura (Punjab), Munir and Bridge [34] reported its presence for the first time in the country and in 2007, Mg was detected in Nepal [35]. The occurrence of Mg in Africa was recorded on grass roots of Paspalum sp. in the South East region of Antsirabe, and its identification was based on morphological traits [36]. Later, in 2014, during a survey carried out in 14 sites distributed along a NW/SE axis between the towns of Marovovay and Manakara, Mg was found [37]. The first report of Mg in South America was by Monteiro et al. [38] in cyperaceas collected in Presidente Prudente, São Paulo, Brazil. However, only in 1991, Sperandio and Monteiro [39] first reported and described the species in the municipality of Palmares do Sul (Rio Grande do Sul) and, in 1994, Sperandio and Amaral [40] found Mg in other municipalities in the south of Rio Grande do Sul. The latest reports confirm the presence of the rice RKN in the region [41,42][41][42]. In Ecuador, Mg was first identified in 1987, in the “Sausalito” village located in the corner of Puerto Inca, province of Guayas, in a field planted with the cultivar Oryzica 1. In surveys conducted in the Provinces of Manabí, Guayas, and Los Ríos, Mg was not found in any other field planted with rice. Nevertheless, by 2000, it had already been disseminated to all rice fields of the Province of Guayas and, in 2002, it was present in the Province of Los Ríos [43]. In a new survey conducted in 2015 in the provinces of Guayas and Los Ríos, the rice RKN was found to be the most widespread, occurring in both rainfed lowland and irrigated areas in high densities [13]. In Colombia, Goméz et al. [44] reported the presence of galls in the roots of rice plants in the county of Tolima, Ibague. Thirteen years later, in a survey programme established by the Colombian rice federation “FEDEARROZ”, Bastidas and Montealegre [45] described the symptoms of a new rice disease denominated as “Entorchamiento” and concluded that it was caused by nematodes of the Meloidogyne genus. The species Mg was later identified, on the basis of morphological and biometrical characters, in other counties and its presence confirmed in other rice production zones, corroborating its spread throughout the country [46,47][46][47]. In Europe, Mg was detected, in July 2016, in several rice fields of northern Italy in the Piedmont region, being the first report of its presence in the EPPO region [14]. Due to this detection, the EPPO decided to include Mg in the Alert List A2 in 2017. Following the first report, it was detected in the Lombardy region, province of Pavia [2]. This Meloidogyne species is present almost in every continent (Table 1. Figure 1). Such occurrence and increase detection draws attention to its potential to affect temperate rice agro-systems adversely.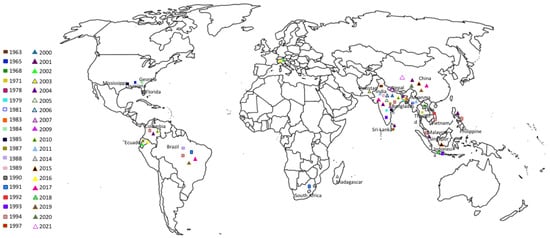
Figure 1. Geographical distribution of Meloidogyne graminicola.
Table 1. Distribution of Meloidogyne graminicola in Africa, America, Asia, and Europe.
| Distribution | Year | References |
|---|
Table 3. Weeds hosts of Meloidogyne graminicola.
| Family | Species (Common Name) | Reference | Family | Species (Common Name) | Reference |
|---|
| Africa | |||||||||
| Madagascar | 2014 | [37] | |||||||
| South Africa | 1991 | [36] | |||||||
| America (North-USA) | |||||||||
| Florida | 2003 | [8] | |||||||
| Georgia | 1984 | [6] | |||||||
| Louisiana | 1965 | [4] | |||||||
| Mississippi | 1990 | [7] | |||||||
| America (South) | |||||||||
| Brazil | 1988, 1991, 1994, 2017, 2019 | [38,39,,48] | [38 | 40, | ][39 | 41, | ][40][41] | 42 | [42][48] |
| Colombia | 1994, 2001, 2010 | [45,46,47] | [45][46][47] | ||||||
| Ecuador | 1987, 2002, 2016 | [13,43] | [13][43] | ||||||
| Asia | |||||||||
| Bangladesh | 1971, 1978, 1979, 1983, 1990 | [49,50,51,52] | [49][50][51][52] | ||||||
| China | 2001, 2015, 2017, 2019, 2020, 2021 | [18,19,20,21,22,53] | [18][19][20][21][22][53] | ||||||
| Indonesia | 1993, 2015, 2018 | [54,55,56] | [54][55][56] | ||||||
| India | 1963, 1979, 1985, 1987, 1989, 1993, 1994, 2000, 2004, 2005, 2006, 2007, 2010, 2011, 2017 |
[3,23,57,58,59,60,61,62,63,64,65,66,67,68,69] | [3][23][57][58][59][60][61][62][63][64][65][66][67][68][69] | ||||||
| Laos | 1968 | [70,71] | [70][71] | ||||||
| Malaysia | 1994 | [72] | |||||||
| Myanmar | 1981, 2011 | [73, | [73 | 74] | ][74] | ||||
| Nepal | 2007, 2009 | [16,35] | [16][35] | ||||||
| Pakistan | 2003 | [34] | |||||||
| Philippines | 1994, 2001 | [75,76] | [75][76] | ||||||
| Singapore | 2001 | [77] | |||||||
| Sri-Lanka | 1997, 2001 | [30,31] | [30][31] | ||||||
| Thailand | 1971 | [25] | |||||||
| Vietnam | 1992, 1994 | [33,78] | [33][78] | ||||||
| Europe | |||||||||
| Italy | 2016, 2018 | [2,14] | [2][14] | ||||||
3. Damage/Crop Losses in Rice
Mg is the most prevalent PPN on rice and considered a major threat to rice as yield losses can reach up to 70% [12,94,112][12][79][80]. Mg densities of 120, 250, and 600 eggs/plant in seedlings 10, 30, and 60 days after planting were reported by Rao et al. [110][81], causing 10% losses. In a later study, Cuc and Prot [65] stated that a density of 100 J2/g root could be considered as high infestation. Most recently, Win et al. [73] found that population densities could exceed 1000 J2/g root with 12–16 galls/plant, contributing to a 65% yield reduction. It has also been found that there is a decline in yield when more than 75% of the roots are affected by nematodes [32]. Additionally, the water regime is an important environmental factor that influences the development and population dynamics of Mg, and the damage and yield loss that it can cause to rice. Soriano et al. [91][82] showed that rice cultivar tolerance to Mg varies with the water regime and that yield losses may be prevented or minimized when the rice crop is flooded early and maintain inundated until harvesting. For example, losses in lowland rainfed rice in Bangladesh can range between 16 and 20%, while in India, losses range between 16 and 32% under irrigated conditions and between 11 and 73% under flooded conditions [102,113][83][84]. In China, the highest incidence of the disease is in the Hunan provinces, exceeding 85% in infested paddy fields [19]. Furthermore, reports of Mg infestations in rice–wheat agroecosystem of India, Nepal, and Pakistan suggest that the damage caused by the rice RKN may be responsible for the poor productivity in this cropping system [10,11,35,114][10][11][35][85].
Changes in agricultural policy and adoption of new rice production technologies in South East Asian countries have influenced the status of the rice RKN problem [64]. For instance, in the Philippines, Mg became a major constrain due to the intensification of rice cropping and shortage of water supply. This situation forced the farmers to grow direct wet seeding, and intermittent irrigation, providing favorable conditions for Mg infestation and increasing the economic losses [9,64][9][64]. In India, the system of rice cultivation shifted to the so-called “system of rice intensification practice”, where a new ecological condition is being developed through modification of rice cultivation practices that includes planting younger and tender seedlings, the creation of greater aeration in soil, and regulation in irrigation. All these conditions provide a suitable environment to increase the infestation levels of the rice RKN [112,114,115][80][85][86].
Spatio-temporal studies have also demonstrated that densities of Mg J2 in the soil fluctuate throughout the year [116][87]. Moreover, Mg’s ability to survive and reproduce in off-seasons on weeds and forage crops contributes to increase the population levels in the soil, and rice infection in the next season [35]. Besides alternative hosts and irrigation, the soil type influenced the tolerance of plants to Mg and showed differences in the multiplication of the nematode [91][82]. Studies have also revealed that infestation levels depend on the rice cultivar [117[88][89],118], and the aggressiveness differs between populations, suggesting intraspecific variability [35,119][35][90]. It was also found that Mg consists of more than one race. In fact, populations from Florida have shown less aggressiveness and difference on the host infection and reproduction patterns than the Asian populations, and populations from Vietnam are not able to reproduce on tomato (Solanum lycopersicum), soy (Glycyne max), or green beans (Phaseolus vulgaris), despite these species being reported as a host of Mg [16,119,120][16][90][91].
4. Host Plants
In addition to the main host, rice, Mg has a wide range of alternative hosts, including cereals and grasses, as well as dicotyledonous plants [15,120,121][15][91][92] (Table 2). Forty-six weeds commonly growing in or around rice fields were assessed for host suitability and were found to be moderate to good hosts of Mg [122][93]. Khan et al. [100][94] reported 17 weed species and, in 2009, Rich et al. [15] reported 24, which supported the survival and multiplication of Mg in the field, acting as a reservoir of nematodes when rice is not present during crop rotations [15] (Table 3). Furthermore, it was believed that Mg caused yield losses only in rice; however, a reduction of the root length of onion (Allium cepa) was observed, with yield losses of 16–35% in the Philippines [76]. In Nepal, India, Pakistan, and Bangladesh, it is considered a threat to wheat crops and to vegetables, such as aubergine (S. melongena), tomato, and okra (Abelmoschus esculentus) [10,122,123,124,125][10][93][95][96][97].Table 2. Cultivated hosts of Meloidogyne graminicola.
| Family | Species (Common Name) | Reference | Family | Species (Common Name) | Reference | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Amaranthaceae | Beta vulgaris | (Beetroot) | [126] | [98] | Musaceae | Musa | sp. (Banana) | [127] | [99] | ||
| Spinacia oleracea | (Spinach) | [12] | M. acuminate | (Dwarf banana) | [128] | [100] | |||||
| Amaryllidaceae | Allium cepa | (Onion) | |||||||||
| ] | |||||||||||
| [ | 76 | ] | Poaceae | Avena sativa | (Oat) | [5] | |||||
| Plantaginaceae | Scoparia dulcis | (Licorice weed) | [ | 122] | [93] | ||||||
| A. tuberosum | (Chive) | [] | [101] | ||||||||
| A. viridis | (Slender amaranth) | ||||||||||
| Hordeum vulgare | (Barley) | [ | 23 | ] | |||||||
| [ | 122] | [93] | |||||||||
| 129 | |||||||||||
| Poaceae | Agropyron repens | (Quack grass) | [ | 100] | [94] | ||||||
| A. fitsulosum | (welsh onion) | [129] | [101] | ||||||||
| Acanthaceae | Rungia parviflora | [128] | [100] | Andropogon | sp. (Beard grass) | [130] | [102] | ||||
| Apiaceae | Coriandrum sativum | (Coriander) | [126] | [98] | Oryza sativa | (Rice) | [5,6] | [5][6] | |||
| ] | |||||||||||
| Alismataceae | Alisma plantago | (Common water- plantain) | [14] | ||||||
| Apiaceae | |||||||||
| Centella asiatica | |||||||||
| (Spade leaf) | [ | 128] | [100] | Alopecurus | sp. (Foxtails) | [120] | [91] | ||
| Asteraceae | Lactuca sativa | (Lettuce) | [12] | ||||||
| Apocynaceae | Catharanthus roseus | (Periwinkle) | [12] | ||||||
| Saccharum officinarum | (Sugarcane) | [ | 12 | ] | |||||
| A. carolinianus | (Carolina foxtail) | [ | 5 | ] | |||||
| Brassicaceae | Brassica oleracea | (Cabbage) | |||||||
| Asteraceae | |||||||||
| [ | 12 | ||||||||
| Ageratum conyzoides | (Billy-goat- weed) | ||||||||
| ] | |||||||||
| [ | 100 | ] | [ | ||||||
| Sorghum bicolor | (Sorghum) | [ | 12 | ] | |||||
| 94 | ] | Brachiaria mutica | (Buffalo grass) | [100] | [94] | ||||
| B. oleracea | var. botrytis (Cauliflower) | [128] | |||||||
| Blumea | sp. | ||||||||
| [ | 100] | ||||||||
| [ | 130] | [102] | |||||||
| Triticum aestivum | (Wheat) | ||||||||
| B. ramosa | (Brown top millet) | ||||||||
| [ | 10 | , | 123] | [10][95] | |||||
| [ | 100 | ] | [94] | ||||||
| Cucurbitaceae | |||||||||
| Eclipta alba | (False Daisy) | [130] | [102] | ||||||
| Phaseolus vulgaris | (Common bean) | [5] | C. annuum (Pepper) | [124][96] | |||||
| Vigna adiate | (Green gram) | [12] | Solanum lycopersicum (Tomato) | [124][96] | |||||
| Oxalidaceae | Oxalis corniculata | [ | 128 | ] | [100] | ||||
| Amaranthaceae | Alternanthera sessilis | (Sessile joy weed) | [100] | [94] | Papillionaceae | Melilotus alba | (Yellow sweet clover) | [23] | |
| Amaranthus spinosus | (Spiny amaranth) | [40 | |||||||
| Cucumis sativus | (Cucumber) | [12] | |||||||
| Bothriochloa intermedia | |||||||||
| [ | 100 | ] | [ | ||||||
| Zea mays | (Maize) | [ | 12 | ] | |||||
| 94 | ] | ||||||||
| Fabaceae | Glycine max | (Soybean) | [122] | [93] | |||||
| E. prostrata | (Eclipta alba) | ||||||||
| Solanaceae | |||||||||
| [ | 131] | [ | |||||||
| Capsicum frutescens | (Chilli) | ||||||||
| 103 | ] | ||||||||
| [ | 130 | ] | |||||||
| Cynodon dactylon | (Bermuda grass) | [ | 126] | [98] | |||||
| [ | 102 | ||||||||
| Grangea ceruanoides | [130] | [102] | Cymbopogon citratus | (Lemon grass) | [128] | [100] | |||
| G. madraspatensis | [130] | [102] | Dactyloctenium aegyptiu | [100] | [94] | ||||
| V. unguiculata | (Cowpea) | [12] | S. melongena (Aubergine) | ||||||
| Sphaeranthus | sp. | ||||||||
| [ | 124 | ] | |||||||
| [ | 126 | ||||||||
| [ | 96 | ] | |||||||
| ] | [98] | D. annulatum | [23] | ||||||
| Malvaceae | Abelmoschus esculentus | (Okra) | [124] | ||||||
| Sphaeranthus senegalensis | [128] | [100][96] | |||||||
| Digitaria filiformis | |||||||||
| (Crab grass) | [ | 126 | ] | [98] | |||||
| Vernonia cinerea | [128] | [100] | D. longifolia | (False couch grass) | [132] | [104] | |||
| Balsaminaceae | Impatiens balsamina | (Garden balsam) | [12] | D. sanguinalis | (Dewgrass) | [100] | [94] | ||
| Brassicaceae | Brassica juncea | (Brown mustard) | [12] | Echinochloa colona | [130] | [102] | |||
| Brassica | sp. | [12] | E. colonum | [4] | |||||
| Caryophyllaceae | Spergula arvensis | (Corn spurry) | [23] | E. crus-galli | (Barnyard grass) | [5] | |||
| Stellaria media | (Chickweed) | [122] | [93] | E. indica | (Goose grass) | [130] | [102] | ||
| Commelinaceae | Cyanotis cucullata | (Roth) | [132] | [104] | E. unioloides | (Chinese love grass) | [132] | [104] | |
| Commelina benghalensis | [132] | [104] | Eleusine coracana | (Finger millet) | [126] | [98] | |||
| Murdannia keisak | (Marsh dew flower) | [14] | Eragrostis tenella | [128] | [100] | ||||
| Compositae | Gnaphalium coarctatum | [133] | [105] | Imperata cylindrica | (Spikegrass) | [128] | [100] | ||
| Cyperaceae | Cyperus brevifolius | (Kyllinga) | [126] | [98] | Ischaemum rugosum | (Saramolla) | [126] | [98] | |
| C. compressus | (Annual sedge) | [105] | [106] | Leersia hexandra | [134] | [107] | |||
| C. difformis | (Variable Flatsedge) | [135] | [108] | Oplismenus compositus | [122] | [93] | |||
| C. imbricatus | [126] | [98] | Poa annua | (Annual bluegrass) | [40] | ||||
| C. odoratus | (Flats edge) | [136] | [109] | Panicum dichotomiflorum | [40] | ||||
| C. pilosus | (Fuzzy flats edge) | [128] | [100] | P. miliaceum | [122] | [93] | |||
| C. procerus | [126] | [98] | P. sumatrense | [128] | [100] | ||||
| C. pulcherrimus | (Elegant s edge) | [126] | [98] | P. repens | [40] | ||||
| C. rotundus | (Purple nutsedge) | [100] | [94] | Paspalum sanguinola | [130] | [102] | |||
| Fimbristylis complanata | [126] | [98] | Paspalum scrobiculatum | [126] | [98] | ||||
| F. dichotoma | [126] | [98] | Pennisetum glaucum | [128] | [100] | ||||
| F. littoralis | (Lesser fimbristylis) | [126] | [98] | P. pedicellatum | [128] | [100] | |||
| F. miliacea | [130] | [102] | P. typhoides | (Pearl millet) | [122] | [93] | |||
| Fuirena ciliaris | [126] | [98] | Scirpus articulatus | [126] | [98] | ||||
| F. glomerata | [126] | [98] | Setaria italica | (Foxtail millet) | [12] | ||||
| Schoenoplectus articulatus | [128] | [100] | Sporobolus diander | [100] | [94] | ||||
| Euphorbiaceae | Chamaesyce hirta | (Asthma herb) | [ | 136 | ] | Polemoniaceae | Phlox drummondii | (phlox) | [12] |
| Phyllanthus urinaria | [130] | [102] | Pontederiaceae | Heteranthera reniformis | [14] | ||||
| Fabaceae | Desmodium triflorum | [122] | [93] | Monochoria vaginalis | [12] | ||||
| Pisum sativum | (Garden pea) | [12] | Portulacaceae | Portulaca oleracea | [122] | [93] | |||
| Trifolium repens | (White clover) | [12] | Solanaceae | Petunia | sp. | [12] | |||
| Trigonella polyceratia | [23] | Physalis minima | [100] | [94] | |||||
| Hydrocharitaceae | Hydrilla | sp. | [132] | [104] | Sida acuta | (Broom grass) | [132] | [104] | |
| Juncaceae | Juncus microcephalus | [ | 137 | ] | Solanum nigrum | [128] | [100] | ||
| Lamiaceae | Leucas lavandulifolia | [128] | [100] | S. sisymbriifolium | [128] | [100] | |||
| Linderniaceae | Bonnaya brachiata | [122,126] | [93][98] | Sphenocleaceae | Sphenoclea zeylanica | [126] | [98] | ||
| Lindernia | sp. | [ | 134 | ] | Ranunculaceae | Ranunculus | sp. (Buttercup) | [105] | [106] |
| Vandellia | sp. | [130] | [102] | Rubiaceae | Borreira articularis | [138] | [110] | ||
| Lythraceae | Ammannia pentandra | [126] | [98] | Hedyotis diffusa | [128] | [100] | |||
| Onagraceae | Jussieua repens | [130] | [102] | ||||||
| Ludwigia adscendens | (Primrose) | [ | 134 | ] | |||||
5. Management
The best strategy for management of Mg is to prevent the movement of plant and soil that in some cases may adhere to machinery or tools. In a recent pest risk analysis for Mg in Italy, it was concluded that the main ways of dispersion of this nematode are likely to be through the movement of infected plants and infested soil, non-host plants that may have grown near areas infested with Mg, and floating roots or plant material in the water [121][92]. Migrant waterbirds, machinery, and travelers were considered a secondary source of entrance. On the other hand, changes in the water regime (intermittent irrigation or water shortages) in many parts of the world are also contributing to the spread and infectivity of the nematode.
To minimize the losses resulting from Mg, management strategies are of extreme importance, and studies have shown that a combination of methods is the best approach to control this nematode in rice fields. The methods that have been applied to control Mg include the use of synthetic nematicides, known as the most efficient strategy, cultural methods, biological agents, and natural nematicides.
Some synthetic nematicides were, recently, strictly regulated or banned from the market, due to the adverse impacts on the environment and human health, reducing the alternatives for RKN control. Cultural methods (fallowing, soil solarization, crop diversification and rotation, etc.) also appeared to have some efficacy. For instance, crop rotation studies with non-host crops, like sweet potato (Ipomoea batatas), cowpea (Vigna unguiculata), sesame (Sesamum indicum), castor (Ricinus communis), sunflower (Helianthus annuus), soybean (Glycine max), turnip (Brassica rapa subsp. rapa), and cauliflower (Brassica oleracea var. botrytis), showed to prevent Mg development [110,132,173][81][104][111]. Nonetheless, none of these practices have gained importance among farmers, because of the high cost and unsatisfactory results. Furthermore, as many weeds found in rice fields are hosts for Mg, serving as nematode reservoirs for the next crops, a weed management programmme must be implemented to maintain a low nematode population in infested fields.
Alternative strategies, such as the “rice field flooding technique”, used by the Italian National Plant Protection Organization (Ministerial Decree of 6 July 2017) to control Mg, had some effect on the nematode population densities. Mg can still propagate under flooding conditions, but the damage induced by this nematode is lower than in shallow intermittently flooded fields [80,174][112][113]. Nevertheless, this method of control also has some limitations, as there are areas where this practice is not applicable due to the soil structure, characterized by a low water retention capacity, or restriction in water use. Another approach explored by Sacchi et al. [174][113] was the use of rice plants as trap crops. Preliminary results indicate that trap cropping for the management of the rice RKN is efficient in most rice-growing areas, especially those with water shortages. However, additional studies are required to establish the most effective number of trap crop cycles that are necessary to reduce Mg population density. Additionally, this technique, in our opinion, could be highly influenced by climate in northern latitudes in order to sow rice in advance and the cost of machinery and water.
The use of biological control agents, such as the fungi Paecilomyces lilacinus, Trichoderma harzianum, T. viride, and other Trichoderma spp.; the bacteria Bacillus subtilis; and the rhizobacterium Pseudomonas fluorescence, have shown promising results against Mg [175,176,177,178][114][115][116][117]. Studies by Amarasinghe and Hemachandra [178][117], in Sri Lanka, revealed that T. viride reduces gall formation and production of egg masses, which represents a potential strategy to be included in integrated pest management programs.
Similarly, the use of essential oils (EOs) has been explored to control RKN, as an alternative to the synthetic nematicides. The nematicidal effects of EO from spices and medicinal plants on RKN have been widely reported. The high effect of Cymbopogon spp. EO (C. martini motia, C. flexuosusand, and C. winterianus) on J2 mortality has been described [179,180,181][118][119][120]. Chavan et al. [182][121] stated that basil (Ocimum basilicum), peppermint (Mentha×piperita), and lemongrass (Cymbopogon citratus) EOs have nematicidal properties against Mg. In order to confirm the efficacy of these EOs, the in vitro tests must be complemented by in vivo soil-based experiments.
Host plant resistance is an environmentally friendly and cost-effective strategy to mitigate damage caused by Mg. A promising alternative for the control of Mg is the screening of germplasm for genotypes that are resistant/tolerant and the development of resistant/tolerant cultivars [80,108,183][112][122][123]. Resistance sources against Mg have been identified in African wild accessions of rice (O. glaberrima and O. longistaminata and O. rufipogon) [184][124], and variability to a certain extent has been perceived [162][125]. Wild accessions that are partially or fully resistant to Mg can therefore act as resistant donors for interspecific crosses with Asian cultivars of rice [184,185][124][126]. Introgression of O. glaberrima into O. sativa has led, for example, to the new rice for Africa, NERICA cultivars [186][127], but the introgression has not been very successful [187][128]. Therefore, natural resistance in O. sativa cultivars is potentially very important. In Asian rice, using the Bala and Azucena mapping population, chromosomes 1, 2, 6, 7, 9, and 11 have been reported as having quantitative trait loci (QTL) for partial resistance to Mg [111][129]. Mapping of Mg resistance on chromosome 10 in Asian rice (cv. Abhishek), using bulk segregant analysis, was reported by Mhatre et al. [188][130]. A hypersensitivity-like reaction to Mg infection found in the Asian rice cv. Zhonghua 11 suggests that resistance to Mg was qualitative rather than quantitative, involving (a) major gene(s) [189][131]. Galeng-Lawilao et al. [190][132] reported the main effect QTL for field resistance in Asian rice on chromosomes 4, 7, and 9 plus two epistatic interactions (between loci on chromosome 3 and 11, and between 4 and 8).
Few studies have used genome-wide association studies (GWASs) as a viable strategy to identify novel QTLs for PPN resistance or susceptibility in different plants [191,192][133][134]. For example, Dimkpa et al. [191][133] confirmed the robustness of GWAS to screen for rice–nematode interactions and identified two resistant accessions (Khao Pahk Maw and LD 24). Studies carried out, in India, by Hada et al. [193][135] allowed the identification of 40 highly resistant accessions. Alternatively, the profiling of the defense response of 36 rice cultivars to Mg infection revealed a variation in the expression of plant defense genes [194][136]. Among all the selected plant defense genes, the expression of mitogen-activated protein kinases (MAPK20), isochorismate synthase genes (ICS1), nonexpressor of pathogenicity expression genes1 (NPR1), phytoalexin-deficient 4 (PAD4), allene oxidase synthase (AOS2), jasmonic acid-inducible rice myb gene (JAMYB), and 1-aminocyclopropane-1-carboxylic acid oxidase (ACO7) was upregulated, possibly providing resistance against Mg. This observation matches the insignificant expression in the susceptible genotypes. These outcomes are significant and can be exploited for breeding purposes.
References
- Kyndt, T.; Fernandez, D.; Gheysen, G. Plant-parasitic nematode infections in rice: Molecular and cellular insights. Annu. Rev. Phytopathol. 2014, 52, 135–153.
- EPPO. 2018—Reporting Service (2018/196): Mg Found in 2018 in 4 Rice Fields in Lombardia Region (Province of Pavia). Available online: https://gd.eppo.int/reporting/article-6390 (accessed on 1 October 2021).
- Israel, P.; Rao, Y.S.; Rao, Y.R.V.J. Investigations on nematodes in rice and rice soils. Oryza 1963, 1, 125–127.
- Golden, A.M.; Birchfield, W. Meloidogyne graminicola (Heteroderidae) a new species of root-knot nematode from grass. Proc. Helminthol. Soc. Wash. 1965, 32, 228–231.
- Birchfield, W. Host parasite relations and host range studies of a new Meloidogyne species in southern USA. Phytopathology 1965, 55, 1359–1361.
- Minton, N.A.; Tucker, E.T.; Golden, A.M. First report of Meloidogyne graminicola in Georgia. Plant Dis. 1987, 71, 376.
- Windham, G.L.; Golden, A.M. First report of Meloidogyne graminicola in Mississippi. Plant Dis. 1990, 74, 1037.
- Handoo, Z.A.; Klassen, W.; Abdul-Baki, A.; Bryan, H.H.; Wang, Q. First record of rice root-nematode (Meloidogyne graminicola) in Florida. J. Nematol. 2003, 35, 342.
- Soriano, I.R.; Reversat, G. Management of Meloidogyne graminicola and yield of upland rice in South-Luzon, Philippines. Nematology 2003, 5, 879–884.
- Padgham, J.L.; Abawi, G.S.; Duxbury, J.M.; Mazid, M.A. Impact of wheat on Meloidogyne graminicola populations in the rice–wheat system of Bangladesh. Nematropica 2004, 34, 183–190.
- Padgham, J.L.; Duxbury, J.M.; Mazid, A.M.; Abawi, G.S.; Hossain, M. Yield loss caused by Meloidogyne graminicola on lowland rainfed rice in Bangladesh. J. Nematol. 2004, 36, 42–48.
- Bridge, J.; Plowright, R.A.; Peng, D. Nematodes Parasites of Rice. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture, 2nd ed.; Luc, M., Sikora, R.A., Bridge, J., Eds.; CABI Publishing: Wallingford, UK, 2005; pp. 87–130.
- Triviño, C.G.; Santillán, D.N.; Velasco, L.V. Plant-Parasitic nematodes associated with rice in Ecuador. Nematropica 2016, 46, 45–53.
- Fanelli, E.; Cotroneo, A.; Carisio, L.; Troccoli, A.; Grosso, S.; Boero, C.; De Luca, F. Detection and molecular characterization of the rice root-knot nematode Meloidogyne graminicola in Italy. Eur. J. Plant Pathol. 2017, 149, 467–476.
- Rich, J.R.; Brito, J.A.; Kaur, R.; Ferrell, J.A. Weed species as hosts of Meloidogyne: A review. Nematropica 2009, 39, 157–185.
- Pokharel, R.R.; Abawi, G.S.; Duxbury, J.M.; Smat, C.D.; Wang, X.; Brito, J.A. Variability and the recognition of two races in Meloidogyne graminicola. Australas. Plant Pathol. 2010, 39, 326–333.
- Upadhyay, V.; Bhardwaj, N.; Neelam, R.; Sajeesh, P.K. Meloidogyne graminicola (Golden and Birchfield) Threat to Rice Production. Res. J. Agric. For. Sci. 2014, 2, 31–36.
- Zhao, H.H.; Liu, W.Z.; Liang, C.; Duan, X.Y. Meloidogyne graminicola, a new record species from China. Acta Phytopathol. Sin. 2001, 5, 184–189.
- Song, Z.Q.; Zhang, D.Y.; Liu, Y.; Cheng, F.X. First report of Meloidogyne graminicola on rice (Oryza sativa) in Hunan Province, China. Plant Dis. 2017, 12, 2153.
- Xie, J.L.; Xu, X.; Yang, F.; Xue, Q.; Peng, Y.L.; Ji, H.L. First report of root-knot nematode, Meloidogyne graminicola, on rice in Sichuan province, Southwest China. Plant Dis. 2019, 103, 2142.
- Ju, Y.; Wu, X.; Tan, G.; Peng, D.; Xu, J.; Qiu, K.; Wu, H. First report of Meloidogyne graminicola on rice in Anhui province, China. Plant Dis. 2020, 105, 512.
- Liu, M.Y.; Liu, J.; Huang, W.; Peng, D. First Report of Meloidogyne graminicola on Rice in Henan Province, China. Plant Dis. 2021.
- Dabur, A.S.; Taya, A.S.; Bajaj, H.K. Life cycle of Meloidogyne graminicola on paddy and its host range studies. Indian J. Nematol. 2004, 34, 80–84.
- Ahamad, F.; Khan, M.R. Status and yield loss assessment of rice root-knot nematode, Meloidogyne graminicola infestation in Siddharthnagar District, Uttar Pradesh. Ann. Plant Prot. Sci.-Indian J. 2018, 26, 360–364.
- Buangsuwon, D.; Tonboonek, P.; Rujirachoon, G.; Braun, A.J.; Taylor, A.L. Nematodes. In Rice Diseases and Pests of Thailand; Rice Protection Research Centre, Rice Department, Ministry of Agriculture: Bangkok, Thailand, 1971; pp. 61–67.
- Hoque, M.O.; Talukdar, M.J. A new disease of rice caused by the nematode Meloidogyne sp. In Proceedings of the Pakistan Science Conference, Peshawar, Pakistan, 27–30 September 1971.
- Bridge, J.; Page, S.L.J. The rice root-knot nematode, Meloidogyne graminicola, on deep water rice (Oryza sativa subsp. indica). Rev. Nematol. 1982, 5, 225–232.
- Miah, S.A.; Shahjahan, A.K.M.; Hossain, M.A.; Sharma, N.R. A survey of rice disease in Bangladesh. Trop. Pest. Manag. 1985, 31, 208–213.
- Padgham, J.L. Impact of The Rice Root-Knot Nematode (Meloidogyne graminicola) on Lowland Rainfed Rice in Northern Western Bangladesh. Ph.D. Thesis, Cornell University, Ithaca, NY, USA, New York, NY, USA, 2003.
- Ekanayake, H.M.R.K.; Toida, Y. Nematode parasites of agricultural crops and their distribution in Sri Lanka. Jpn. Agric. Res 1997, 4, 23–39.
- Ekanayake, H.M.R.K. Histopathological changes caused by Meloidogyne graminicola in rice roots. Ann. Sri Lanka Dep. Agric. 2001, 3, 43–46.
- Nugaliyadde, L.; Dissanayake, D.M.N.; Herath, H.M.D.N.; Dharmasena, C.M.D.; Jayasundara, D.M. Outbreak of rice root-knot nematode, Meloidogyne graminicola (Golden & Birchfield) in Nikewaratiya, Kurunegala in Maha 2000/2001. (Short Communication). Ann. Sri Lanka Dep. Agric. 2001, 3, 373–374.
- Cuc, N.T.T.; Prot, J. Root-parasitic nematodes of deep-water rice in the Mekong delta of Vietnam. Fundam. Appl. Nematol. 1992, 15, 575–577.
- Munir, A.; Bridge, J. Rice root-knot nematode, Meloidogyne graminicola Golden and Birchfield, 1965 from rice in Pakistan. Pak. J. Nematol. 2003, 21, 133–136.
- Pokharel, R.R.; Abawi, G.S.; Zhang, N.; Duxbury, J.M.; Smart, C.D. Characterization of isolates of Meloidogyne from rice–wheat production fields in Nepal. J. Nematol. 2007, 39, 221–230.
- Kleynhans, K.P. The Root-Knot Nematodes of South Africa; No. 231:61; Technical Communication, Department of Agricultural Development: Pretoria, South Africa, 1991; p. 136.
- Chapuis, E.; Besnard, G.; Andrianasetra, S.; Rakotomalala, M.; Nguyen, H.T.; Bellafiore, S. First report of the root-knot nematode (Meloidogyne graminicola) in Madagascar rice fields. Australas. Plant Dis. Notes 2016, 11, 32.
- Monteiro, A.R.; Ferraz, L.C.C.B. Encontro de Meloidogyne graminicola e primeiro ensaio de hospedabilidade no Brasil. Nematol. Bras 1988, 12, 149–150.
- Sperandio, C.A.; Monteiro, A.R. Ocorrência de Meloidogyne graminicola em arroz irrigado no Rio Grande do Sul. Nematol. Bras. 1991, 15, 24.
- Sperandio, C.A.; Amaral, A.S. Ocorrência de Meloidogyne graminicola causador da falsa bicheira do arroz irrigado no Rio Grande do Sul. Rev. Lavoura Arrozeira 1994, 47, 18–21.
- Bellé, C.; Balardin, R.R.; Dalla Nora, D.; Schmitt, J.; Gabriel, M.; Ramos, R.F.; Antoniolli, Z.I. First Report of Meloidogyne graminicola (Nematoda: Meloidogynidae) on barley (Hordeum vulgare) in Brazil. Plant Dis. 2019, 103, 1045.
- Bellé, C.; Ramos, R.F.; Balardin, R.R.; Kaspary, T.E.; Brida, A.L. Reaction of rice cultivars to Meloidogyne graminicola as a function of irrigation management. Commun. Plant Sci. 2019, 9, 124–128.
- Triviño, C.G.; Velasco, L.V. Problemas que Afectan la Producción de Arroz; EC, 8, 17; Revista Informativa; INIAP (Instituto Nacional Autónomo de Investigación Agropecuarias): Quito, Ecuador, 2013.
- Gómez, J.; Puerta, F.; Gómez, R. Nematodos fitoparásitos asociados a las siembras de arroz en la terraza de Ibagué, Tolima-Colombia. Rev. Arroz 1981, 30, 17–24.
- Bastidas, H.; Montealegre, S.F.A. Aspectos generales de la nueva enfermedad del arroz llamada entorchamiento. Arroz 1994, 43, 392–416.
- Jaraba-Navas, J.; Lozano, Z.; Pérez, C.R. Identificación del nematodo del nudo radical del arroz en los departamentos de Cesar y Guajira. Fitopatol. Colomb. 2001, 25, 29–32.
- Hoyos, L.M.; Moya, J.G. Nematodes associated with rice crops in Huila and Tolima. Agron. Colomb. 2010, 28, 577–579.
- Negretti, R.R.R.D.; Mattos, V.S.; Manica-Berto, R.; Gomes, C.B.; Carneiro, R.M.D.G.; Somavilla, L.; Castagnone-Sereno, P.; Regina, M.D.G. Characterisation of Meloidogyne species complex parasitising rice in southern Brazil. Nematology 2017, 19, 403–412.
- Page, S.L.; Bridge, J.; CABI. Plant nematodes on deep-water rice in Bangladesh. (ODM Report on visit to Bangladesh, 19 June–9 August, 1978). Available online: https://www.cabi.org/isc/abstract/19810881735 (accessed on 1 October 2021).
- Page, S.L.J.; Bridge, J.; Cox, P.; Rahman, L. Root and soil parasitic nematodes of deep water rice areas in Bangladesh. Int. Rice Res. Notes 1979, 4, 10–11.
- Rahman, M.L.; Taylor, B. Nematode pests associated with deep water rice in Bangladesh. Int. Rice Res. Notes 1983, 8, 20–21.
- Rahman, M.L.; Evans, A.A.F.; Miah, S.A. Plant damage and yield loss caused by the rice root-knot nematode, Meloidogyne graminicola in deepwater rice in Bangladesh. Bangladesh J. Bot. 1990, 19, 107–116.
- Zhou, X.; Liu, G.K.; Xiao, S.; Zhang, S. First report of Meloidogyne graminicola infecting banana in China. Plant Dis. 2015, 99, 420.
- Netscher, C.; Erlan, S. A root-knot nematode, Meloidogyne graminicola, parasitic on rice in Indonesia. Afro-Asian J. Nematol. 1993, 3, 90–95.
- Nurjayadi, M.Y.; Munif, A.; Suastika, G. Identification of the root-knot nematode, Meloidogyne graminicola, on rice plants in West. Java. J. Phytopathol. Indones. 2015, 11, 113.
- Mirsam, H.; Kurniawati, F. First report in South Sulawesi: Morphological and molecular characters of root-knot nematodes associated with rice roots in Wajo District, South Sulawesi. Indones. J. Plant Prot. 2018, 22, 58.
- Prasad, K.S.K.; Krishnappa, K.; Rao, Y.S. Response of some improved rice varieties to the development and reproduction of Meloidogyne graminicola Golden and Birchfield, 1965. Mysore J. Agric. Sci. 1979, 13, 60–62.
- Prasad, J.S.; Panwar, M.S.; Rao, Y.S. Occurrence of root knot-nematode, Meloidogyne graminicola in semideepwater rice. Curr. Sci. 1985, 54, 387–388.
- Prasad, J.S.; Panwar, M.S.; Rao, Y.S. Nematode problems of rice in India. Trop. Pest Manag. 1987, 33, 127–136.
- Kaul, V.K.; Chhabra, H.K. A new record of Meloidogyne graminicola on Echinochloa crus-galli in Punjab. Indian J. Nematol. 1989, 19, 76–78.
- Gaur, H.S.; Khan, E.; Sehgal, M. Occurrence of two species of root-knot nematodes infecting rice, wheat and monocot weeds in northern India. Ann. Plant Prot. Sci. 1993, 1, 41–142.
- Nath, R.C.; Mukherjee, B.; Dasgupta, M.K.; Siddiqi, M.R. Prevalence and distribution of plant parasitic nematodes in rice fields of Tripura, India. Afro-Asian J. Nematol. 1994, 4, 147–150.
- Baqri, Q.H.; Ahmad, N. Qualitative and quantitative studies of plant and soil inhabiting nematodes associated with rice crop in Sikkim, India. Rec. Zool. Surv. India 2000, 98, 137–148.
- Sheela, M.S.; Jiji, T.; Nisha, M.S.; Rajkumar, J. A new record of Meloidogyne graminicola on rice, Oryza sativa L in Kerala. Indian J. Nematol. 2005, 35, 218.
- Prasad, J.S.; Vishakanta, L.; Gubbaiah, V. Outbreak of root-knot nematode (Meloidogyne graminicola) disease in rice and farmers perceptions. Indian J. Nematol. 2006, 36, 85–88.
- Singh, V.K.; Kalia, C.S.; Kaul, V. New record of root-knot nematode, Meloidogyne graminicola infecting rice in Jammu. Indian J. Nematol. 2007, 37, 94.
- Prasad, J.S.; Somasekhar, N.; Varaprasad, K.S. Nematode infestation in paddy. In Nematode Infestations; Part I: Food Crop; Khan, M.R., Jairajpuri, M.S., Eds.; Indian Academy of Sciences: Bengaluru, India, 2010; pp. 17–71.
- Pankaj, A.S.; Jain, R.K.; Singh, K. Incidence of Meloidogyne graminicola on rice in Andaman Islands. Ann. Plant Prot. Sci. 2011, 19, 259–260.
- Salalia, R.; Walia, R.K.; Somvanshi, V.S.; Kumar, P.; Kumar, A. Morphological, morphometric, and molecular characterization of intraspecifc variations within Indian populations of Meloidogyne graminicola. J. Nematol. 2017, 49, 254–267.
- Manser, P.D. Meloidogyne graminicola a cause of root-knot of rice. FAO Plant Prot. Bull. 1968, 16, 1–11.
- Manser, P.D. Notes on the rice root-knot nematode in Laos. FAO Plant Prot. Bull. 1971, 19, 138–139.
- Zainal-Abidin, A.A.; Monen-Abdullah, M.A.; Azawiyah, A.H. Meloidogyne graminicola: A new threat to rice cultivation in Malaysia. In Proceedings of the 4th International Conference on Plant Protection in the Tropics, Kuala Lumpur, Malaysia, 28–31 March 1994; pp. 246–247.
- Myint, Y.Y. Country report on root-knot nematode in Burma. In Proceedings of the 3rd Research Planning Conference on Root-knot Nematodes, Meloidogyne spp., Region VI, Raleigh, NC, USA, 20–24 July 1981; North Carolina State University: Jakarta, Indonesia; pp. 163–170.
- Win, P.P.; Kyi, P.P.; DeWaele, D. Effect of agro-ecosystem on the occurrence of the rice root-knot nematode Meloidogyne graminicola on rice in Myanmar. Australas. Plant Pathol. 2011, 40, 187–196.
- Prot, J.C.; Soriano, I.R.S.; Matias, D. Major root-parasitic nematodes associated with irrigated rice in the Philippines. Fundam. Appl. Nematol. 1994, 17, 75–78.
- Gergon, E.B.; Miller, S.A.; Halbrendt, J.M.; Davide, R.G. Effect of rice root-knot nematode on growth and yield of Yellow Granex scallion. Plant Dis. 2002, 86, 1339–1344.
- AVA. Diagnostic Records of the Plant Health Diagnostic Services; Plant Health Centre, Agri-Food & Veterinary Authority: Singapore, 2001.
- Cuc, T.T.; Prot, J.C. Nematode parasites of deepwater and irrigated rice in the Mekong River Delta. In Proceedings of the conference held by Vietnam and IRRI: A Partnership in Rice Research, Hanoi, Vietnam, 4–7 May 1994; pp. 51–260.
- Plowright, R.; Bridge, J. Effect of Meloidogyne graminicola (Nematoda) on the establishment, growth and yield of rice cv.IR36. Nematology 1990, 36, 81–89.
- Khan, M.R.; Jain, R.K.; Ghule, T.M.; Pal, S. Root-knot Nematodes in India-A Comprehensive Monograph. In All India Coordinated Research Project on Plant Parasitic Nematodes with Integrated Approach for their Control; Indian Agricultural Research Institute: New Delhi, India, 2014.
- Rao, Y.S.; Prasad, J.S.; Yadava, C.P.; Padalia, C.R. Influence of rotation crops in rice soils on the dynamics of parasitic nematodes. Biol. Agric. Hortic. 1984, 2, 69–78.
- Soriano, I.R.S.; Prot, J.C.; Matias, D.M. Expression of tolerance for Meloidogyne graminicola in rice cultivars as affected by soil type and flooding. J. Nematol. 2000, 32, 309–317.
- Tian, Z.L.; Maria, M.; Barsalote, E.M.; Castillo, P.; Zheng, J.W. Morphological and molecular characterization of the rice root-knot nematode, Meloidogyne graminicola, Golden and Birchfield, 1965 occurring in Zhejiang, China. J. Integr. Agric. 2018, 17, 2724–2733.
- Prasad, J.S.; Panwar, M.S.; Rao, Y.S. Screening of some rice cultivars against the root-knot nematode Meloidogyne graminicola. Indian J. Nematol. 1986, 16, 112–113.
- Jain, R.K.; Khan, M.R.; Kumar, V. Rice root-knot nematode (Meloidogyne graminicola) infestation in rice. Arch. Phytopathol. Plant Prot. 2012, 45, 635–645.
- Haque, Z.; Khan, M.R.; Ahamad, F. Relative antagonistic potential of some rhizosphere biocontrol agents for the management of rice root-knot nematode, Meloidogyne graminicola. Biol. Control 2018, 126, 109–116.
- Win, P.P.; Kyi, P.P.; Maung, Z.T.Z.; De Waele, D. Population dynamics of Meloidogyne graminicola and Hirschmanniella oryzae in a double rice-cropping sequence in the lowlands of Myanmar. Nematology 2013, 15, 795–807.
- Amarasinghe, L.D.; Kariyapperuma, K.A.D.P.S.; Pathirana, H.N.I. Study on approaches to integrated control of Meloidogyne graminicola in rice. J. Sci. Univ. Kelaniya 2007, 3, 29–46.
- Amarasinghe, L.D. An integrated approached to the management of rice root-knot nematode, Meloidogyne graminicola in Sri Lanka. J. Sci. Univ. Kelaniya 2011, 6, 55–63.
- Bellafiore, S.; Jougla, C.; Chapuis, E.; Besnard, G.; Suong, M.; Vu, P.N.; De Waele, D.; Gantet, P.; Thi, X.N. Intraspecific variability of the facultative meiotic parthenogenetic root-knot nematode (Meloidogyne graminicola) from rice fields in Vietnam. Comptes Rendus Biol. 2015, 338, 471–483.
- McGowan, J.B.; Langdon, K.R. Host of the rice root-knot nematode Meloidogyne graminicola. In Nematology Circular, No. 172; Florida Department of Agriculture: Gainesville, FL, USA, 1989.
- Torrini, G.; Roversi, P.F.; Cesaroni, C.F.; Marianelli, L. Pest risk analysis of rice root-knot nematode (Meloidogyne graminicola) for the Italian territory. EPPO Bull. 2020, 50, 330–339.
- Roy, A.K. Host suitability of some crops to Meloidogyne graminicola. Indian J. Nematol. 1977, 30, 483–485.
- Khan, M.R.; Ghosh, S.; Bhattacharya, S.P. Weed hosts of rice root-knot nematode, Meloidogyne graminicola from West Bengal. Ecol. Environ. 2004, 22, 583–584.
- Soomro, M.H.; Hauge, N.G.M. Relationship between inoculum density of Meloidogyne graminicola, growth of rice seedling and development of the nematode. Pak. J. Nematol. 1992, 11, 103–114.
- Duxbury, J.M. Sustainability of Post-Green Revolution Agriculture: The Rice-Wheat Cropping System of South Asia; Annual Report; Soil Management CRSP Management Entity, University of Hawaii: Honolulu, HI, USA, 2001.
- Pokharel, R.R.; Abawi, G.S.; Duxbury, J.M.; Smart, C. Characterization of root-knot nematodes recovered from rice-wheat fields in Nepal. J. Nematol. 2004, 36, 341–342.
- Usha, D.; Khetarpal, R.K.; Agarwal, P.C.; Ijun Lal, A.; Manju, L.K.; Gupta, K.; Parak, D.B. Potential Quarantine Pests for India: Cereals; NBPGR: New Delhi, India, 2005.
- Reversat, G.; Soriano, I. The potential role of bananas in spreading rice root-knot nematode, Meloidogyne graminicola. Int. Rice Res. Notes 2002, 27, 23–24.
- EPPO 2016–Reporting Service (2016/211): First report of Meloidogyne graminicola in Italy. Available online: https://gd.eppo.int/reporting/article-5956 (accessed on 1 November 2021).
- Chen, J.W.; Chen, S.Y.; Ning, X.L.; Shi, C.H.; Cheng, X.; Xiao, S.; Liu, G.K. First report of Meloidogyne graminicola infecting Chinese chive in China. Plant Dis. 2019, 103, 2967.
- Rao, Y.S.; Israel, P.; Biswas, H. Weed and rotation crop plants as hosts for the rice root-knot nematode, Meloidogyne graminicola (Golden and Birchfield). Oryza 1970, 7, 137–142.
- Brito, J.A.; Kaur, R.; Cetintas, R.; Stanley, J.D.; Mendes, M.L.; McAvoy, E.J.; Powers, T.O.; Dickson, D.W. Identification and isozyme characterization of Meloidogyne spp. infecting horticultural and agronomic crops, and weed plants in Florida. Nematology 2008, 10, 757–766.
- Ravindra, H.; Sehgal, M.; Narasimhamurthy, H.B.; Jayalakshmi, K.; Khan, I. Rice root-knot nematode (Meloidogyne graminicola) an emerging problem. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 3143–3171.
- Belair, G.; Benoit, D.L. Host susceptibility of 32 common weeds to Meloidogyne hapla in organic soils of southwestern Quebec. J. Nematol. 1996, 28, 643–647.
- Yik, C.P.; Birchfield, W. Host studies and reactions of cultivars to Meloidogyne graminicola. Phytopathology 1979, 69, 497–499.
- Mulyadi, M.; Triman, B. Study on host plant of root-knot nematode of rice. Indonesian J. Plant Prot. 1995, 1, 8–11.
- Bajaj, H.K.; Dabur, K.R. Cyperus deformis, a new host record of rice root-knot nematode, Meloidogyne graminicola. Indian J. Nematol. 2000, 30, 25.
- Mani, A.; Hinai, M.A. Host range and distribution of Meloidogyne incognita and M. javanica in the Sultanate of Oman. Nematropica 1996, 26, 73–79.
- Gowda, D.N.; Kurdikeri, C.B.; Gowda, C.K. Weeds as hosts of root-knot nematodes. Indian J. Nematol. 1995, 25, 215–216.
- Rao, Y.S. Research on rice nematodes. In Rice in India; Padmanabhan, S.Y., Ed.; ICAR Monograph: New Delhi, India, 1985; pp. 591–615.
- Mantelin, S.; Bellafiore, S.; Kyndt, T. Meloidogyne graminicola: A major threat to rice agriculture. Mol. Plant Pathol. 2017, 18, 3–15.
- Sacchi, S.; Torrini, G.; Marianelli, L.; Mazza, G.; Fumagalli, A.; Cavagna, B.; Ciampitti, M.; Roversi, P.F. Control of Meloidogyne graminicola a root-knot nematode using rice plants as trap crops: Preliminary results. Agriculture 2021, 11, 37.
- Huong, T.T.L.; Padgham, J.L.; Sikora, R.A. Biological control of the rice root-knot nematode Meloidogyne graminicola on rice, using endophytic and rhizosphere fungi. Int. J. Pest Manag. 2009, 55, 31–36.
- Narasimhamurthy, H.B.; Ravindra, H.; Mukesh, S.; Ekabote, S.D.; Ganapathi, G. Bio-management of rice root-knot nematode (Meloidogyne graminicola). J. Entomol. Zool. Stud. 2017, 5, 1433–1439.
- Seenivasan, N.; David, P.M.M.; Vivekanandan, P.; Samiappan, R. Biological control of rice root-knot nematode, Meloidogyne graminicola, through mixture of Pseudomonas fluorescens strains. Biocontrol Sci. Technol. 2021, 22, 611–632.
- Amarasinghe, L.D.; Hemachandra, K.H.D.J.K. Meloidogyne graminicola infestation in selected Sri Lankan rice varieties, Oryza sativa L. and nemato-toxic effect of Trichoderma viride to reduce infectivity. J. Sci. Univ. Kelaniya 2020, 13, 18–34.
- Sangwan, N.K.; Verma, K.K.; Verma, B.S.; Malik, M.S.; Dhindsa, K.S. Nematicidal activity of EOs of Cymbopogon grasses. Nematologica 1985, 31, 93–99.
- Saxena, D.B.; Goswami, B.K.; Tomar, S.S. Nematicidal activity of some EOs against Meloidogyne incogn. Indian Perfum. 1987, 31, 150–154.
- Oka, Y.; Nacar, S.; Putievsky, E.; Ravid, U.; Yaniv, Z.; Spiegel, Y. Nematicidal activity of EOs and their components against the root-knot nematode. Phytopathology 2000, 90, 710–715.
- Chavan, S.N.; Somasekhar, N.; Rani, J. Nematicidal activity of essential oils against rice root-knot nematode Meloidogyne graminicola. Indian J. Nematol. 2019, 49, 135–141.
- Dutta, T.K.; Ganguly, A.K.; Gaur, H.S. Global status of rice root-knot nematode, Meloidogyne graminicola. Afr. J. Microbiol. Res. 2012, 6, 6016–6021.
- Pokharel, R.R.; Duxbury, J.M.; Abawai, G. Evaluation of Protocol for Assessing the Reaction of Rice and Wheat Germplasm to Infection by Meloidogyne graminicola. J. Nematol. 2012, 44, 274–283.
- Soriano, I.; Schmit, V.; Brar, D.; Prot, J.C.; Reversat, G. Resistance to rice root-knot nematode Meloidogyne graminicola identified in Oryza longistaminata and O. glaberrima. Nematology 1999, 1, 95–398.
- Kumari, C.; Dutta, T.K.; Banakar, P.; Rao, U. Comparing the defence-related gene expression changes upon root-knot nematode attack in susceptible versus resistant cultivars of rice. Sci. Rep. 2016, 6, 22846.
- Plowright, R.; Coyne, D.L.; Nash, P.; Jones, M.P. Resistance to the rice nematodes Heterodera sacchari, Meloidogyne graminicola and M. incognita in Oryza glaberrima and O. glaberrima x O. sativa interspecies hybrids. Nematology 1999, 1, 745–751.
- Dibba, L.; Zeller, M.; Diagne, A. The impact of new Rice for Africa (NERICA) adoption on household food security and health in the Gambia. Food Secur. 2017, 9, 929–944.
- Cabasan, M.T.N.; Kumar, A.; De Waele, D. Evaluation of resistance and tolerance of rice genotypes from crosses of Oryza glaberrima and O. sativa to the rice root-knot nematode, Meloidogyne graminicola. Trop. Plant Pathol. 2017, 43, 230–241.
- Shrestha, R.; Uzzo, F.; Wilson, M.J.; Price, A.H. Physiological and genetic mapping study of tolerance to root-knot nematode in rice. New Phytol. 2007, 3, 665–672.
- Mhatre, P.H.; Pankaj, A.S.; Singh, A.K.; Ellur, R.K.; Kumar, P. Molecular mapping of rice root-knot nematode (Meloidogyne graminicola) resistance gene in Asian rice (Oryza sativa L.) using STMS markers. Indian J. Genet. 2017, 77, 163–165.
- Phan, N.T.; De Waele, D.; Lorieux, M.; Xiong, L.; Bellafiore, S. A hypersensitivity-like response to Meloidogyne graminicola in rice (Oryza sativa L.). Phytopathology 2018, 108, 521–528.
- Galeng-Lawilao, J.; Kumar, A.; De Waele, D. QTL mapping for resistance to and tolerance for the rice root-knot nematode, Meloidogyne graminicola. BMC Genet. 2018, 19, 53.
- Dimkpa, S.O.N.; Lahari, Z.; Shrestha, R.; Douglas, A.; Gheysen, G.; Price, A.H. A genome-wide association study of a global rice panel reveals resistance in Oryza sativa to root-knot nematodes. J. Exp. Bot. 2016, 67, 1191–1200.
- Warmerdam, S.; Sterken, M.G.; Van Schaik, C.; Oortwijn, M.E.; Sukarta, O.C.; Lozano-Torres, J.L.; Smant, G. Genome-wide association mapping of the architecture of susceptibility to the root-knot nematode Meloidogyne incognita in Arabidiopsis thaliana. New Phytol. 2018, 218, 724–737.
- Hada, A.; Dutta, T.K.; Singh, N.; Singh, B.; Rai, V.; Singh, N.K.; Rao, U. A genome-wide association study in Indian wild rice accessions for resistance to the root-knot nematode Meloidogyne graminicola. PLoS ONE 2020, 15, e0239085.
- Hatzade, B.; Singh, D.; Phani, V.; Kumbhar, S.; Rao, U. Profiling of defence responsive pathway regulatory genes in Asian rice (Oryza sativa L.) against infection of Meloidogyne graminicola (Nematoda: Meloidogynidae). 3 Biotech 2020, 10, 60.
More