Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Woan-Yuh Tarn and Version 2 by Lindsay Dong.
Antisense oligonucleotide (ASO) technology exploits a single-strand short oligonucleotide to either cause target RNA degradation or sterically block the binding of cellular factors or machineries to the target RNA. Chemical modification or bioconjugation of ASOs can enhance both its pharmacokinetic and pharmacodynamic performance, and it enables customization for a specific clinical purpose. ASO-based therapies have been used for treatment of genetic disorders, cancer and viral infections. In particular, ASOs can be rapidly developed for newly emerging virus and their reemerging variants.
- virus
- antisense oligonucleotide
1. Development and Delivery of Therapeutic ASOs
ASO is a single-stranded DNA of 12–25 nucleotides in length targeting diseases-associated transcripts [1][2][3][4][13,14,15,16]. The first ASO that inhibits the replication of Rous sarcoma virus and cell transformation was reported in 1978 [5][17]. ASOs modulate target gene expression via one of the following mechanisms [1][2][3][4][13,14,15,16]. (1) ASO forms a hybrid with the target RNA, inducing RNA cleavage by RNase H, which is an endonuclease cleaving the RNA strand of a DNA-RNA duplex. (2) ASO base-pairs with a functional cis-element of the target RNA and hence blocks access of cellular machinery to the RNA. Such steric-blocking ASOs may cause splice isoform switch or translation inhibition or may alter the stability of the target RNA (Figure 1).
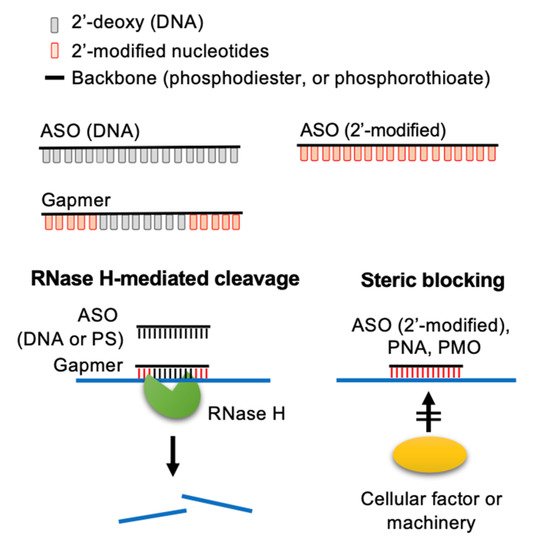
Figure 1. Molecular mechanisms of action of ASOs. ASOs can modulate the expression of target RNAs via two different mechanisms. Conventionally, ASOs cause RNase H-mediated cleavage of the target RNA. Additionally, 2′-O-modified ASOs and neutral DNA mimics (PMOs and PNAs) act as a steric-blocker to prevent the access of cellular factors to the target RNA. Adapted from [3], Springer Nature Limited, 2020.
1.1. Modifications of ASOs
1.1. Modifications of ASOs
Chemical modifications have been developed to improve the pharmacokinetic properties of ASOs, including stability, specificity, and membrane permeability, and minimize their cytotoxicity. Three generations of ASOs have been broadly classified with respect to the types of modifications [6][7][8][18,19,20]. In the first-generation ASOs, one of the non-bridging oxygen atoms in the phosphodiester bond are replaced, resulting in a phosphoramidate, methylphosphonate, or phosphorothioate (PS) linkage (Figure 2A, PS). The PS linkage improves membrane penetration of ASOs and does not interfere with RNase H-mediated RNA cleavage [6][7][8][18,19,20]. PS-ASOs can bind proteins in plasma, thereby preventing rapid clearance of the ASOs from the circulatory system [9][10][21,22]. PS chirality (Rp and Sp) may influence the pharmacological properties of ASOs, such as the stability and RNase H1 cleavage patterns [11][12][23,24]. The second-generation ASOs are modified with an alkyl moiety, such as a methyl (2′-OMe) or methoxyethyl (2′-MOE) group at the 2′ position of the ribose (Figure 2A). These ASOs have greater affinity for their target RNAs and lesser cytotoxicity but cannot recruit RNase H1 for RNA cleavage [13][14][25,26]. The third-generation ASOs include locked nucleic acid (LNA), peptide nucleic acid (PNA), and phosphorodiamidate morpholino oligomer (PMO) (Figure 2A).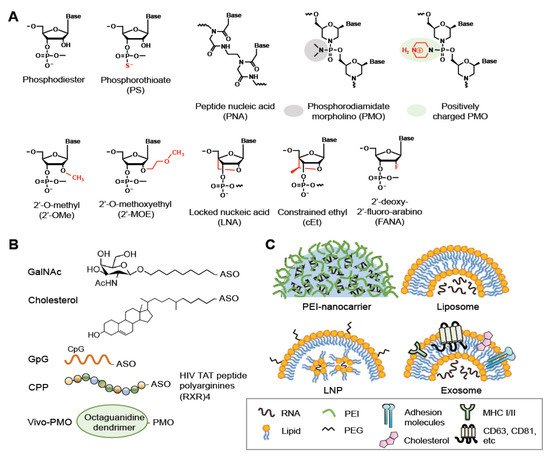
Figure 2. Modifications, bioconjugations and delivery vehicles of ASOs. (A) Structures of backbone or sugar-modified ASOs as well as PNA and PMO oligomers. (B) Bioconjugates of ASOs include GalNac, Cholesterol, CpG DNA and CPP (R, arginine; X, 6-aminohexanoic acid). Vivo-PMO is an PMO covalently linked to an octa-guanidine dendrimer. (C) Representative delivery vehicles of ASOs include EPI-nanocarrier, liposome, LNP, and exosome. Adapted from [3], Spring Nature Limited, 2020; [8], MDPI, 2021.
1.2. Bioconjugations of ASOs
Nucleic acid-based drugs generally enter cells via endocytosis; most of these therapeutics must be formulated as a bioconjugate to facilitate receptor-mediated endocytosis and/or increase their lipophilicity [19][20][21][36,37,38] (Figure 2B). For example, conjugation of an siRNA with anandamide, folate, or cholesterol enables efficient uptake of the siRNA by cells, likely via the corresponding conjugate-specific receptor [22][39]. Notably, a PS backbone assists the membrane translocation of ASOs; thus, bioconjugation and/or delivery agents are often dispensable for PS-ASOs [9][10][21,22]. Therapeutic oligonucleotides also can be conjugated to certain ligands that bind cell type-specific receptors. For example, glycoproteins terminating with N-acetylgalactosamine (GalNAc) can be recognized by the asialoglycoprotein receptor (ASGPR), which exists primarily on the cell-surface of hepatocytes [23][40]. Therefore, conjugation with GalNAc can promote the uptake of siRNAs and ASOs by hepatocytes via ASGPR-mediated endocytosis. GalNAc-2′-MOE-ASOs targeting the mRNA encoding the liver glucagon receptor have been designed for treatment of type 2 diabetes [24][41]. CpG dinucleotides can lead to the uptake of conjugated oligonucleotides by dendritic cells or macrophages that express innate immune receptors [25][42]. Besides the aforementioned conjugates, a number of cell-penetrating peptides (CPPs) have been developed to enhance drug delivery, including polycationic HIV-1 Tat peptide, the hydrophobic residue-containing peptide penetratin that is derived from the Drosophila antennapedia homeodomain, and artificial poly-arginine peptides [26][43]. CPPs may undergo endocytosis or directly penetrate cells [26][43]. Composite CPPs consisting of penetratin and 6-aminohexanoic-spaced oligo-arginine (RXR) have been used to enhance the efficiency of delivery of charge-neutral PMOs or PNAs to cells in vivo or in culture [27][44]. Various conditionally activatable CPPs have been designed for selective delivery. For example, a pH-sensitive transportan CPP bearing lysine-to-histidine substitutions can enter cells under acidic conditions, such as the tumor microenvironment [28][45]. Finally, because guanidinium groups of arginine-rich peptides are critical for peptide translocation across the plasma membrane, a synthetic octa-guanidine dendrimer has been conjugated to PMOs, and such conjugates are called vivo-PMOs [29][46]. Vivo-PMOs, although widely used for transient gene silencing in vitro, cause coagulation owing to dendrimer clustering in animals; supplementation of Vivo-PMO with anticoagulants may counteract its toxicity [30][47].1.3. Vehicle-Mediated Delivery of ASOs
Oligonucleotide bioconjugates offer the potential for enhanced drug delivery, but recent advances in nanotechnology have further benefited the transport of therapeutic ASOs across biological barriers and improved their pharmacokinetics in circulating blood. Several nanoparticle-mediated delivery systems have been developed [3][6][19][20][21][15,18,36,37,38] (Figure 2C). Cationic polymer nanocarriers are formed via ionic interactions between negatively charged ASOs and positively charged macromolecules such as polyethylenimine (PEI). PEI promotes cellular delivery of ASOs but is somewhat cytotoxic. Modification with phospholipid (such as dioleoylphosphatidylethanolamine, DOPE) or copolymerization with polyethylene glycol (PEG) can enhance the efficiency of PEI in ASO delivery and reduce its cytotoxicity [31][48]. Moreover, bioconjugation with cell-binding ligands such as transferrin, antibodies, or carbohydrates can facilitate receptor-mediated uptake of nanocarriers [6][18]. LNPs are nanoparticles mainly constructed with lipids. Among them, liposomes are spherical vesicles comprising single or multiple lipid bilayers. ASOs can be carried in the aqueous space encapsulated by artificial liposomes. Most liposomes formulated for RNA delivery comprise both cationic lipids and neutral lipids such as DOPE; such a lipid combination enhances the transfection efficiency and reduces the cytotoxicity of liposomes [32][49]. At present, LNPs represent a highly potent RNA delivery vehicle; for such LNPs, nucleic acids are organized in inverse lipid micelles inside the nanoparticle [33][50]. Exosomes are naturally secreted extracellular vesicles that transfer macromolecules between cells [34][53]. For drug delivery, exosomes have both advantages and drawbacks. For example, they have inherent anti-inflammatory properties, can traverse biological membranes such as the blood-brain barrier, and can be produced in an autologous manner, but they are heterogenous and uneasy for large-scale production [35][54]. Exosomes can be modified through chemical methods or genetic engineering. Fusion of green fluorescent protein to the exosome surface protein CD63 allows tracking of the exosome and monitoring of cargo delivery [36][55]. Coating of exosomes with cationic lipids and a pH-sensitive amphipathic peptide can enhance cellular uptake and fusion with endosomes and subsequent cargo release [37][56].23. ASOs Targeting Viruses
The development of more efficacious treatments against various viral diseases from acute to persistent infection is still in high demand. Among different nucleic acid-based therapies, ASOs directly act on viral genomic RNA or transcripts (Figure 3) and can be rationally designed for any new virus (or variants) or a reemergent virus.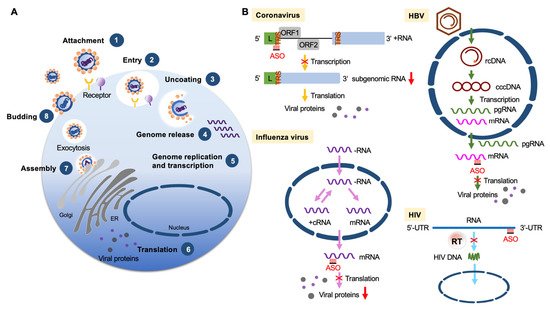
Figure 3. ASOs targeting viruses. (A) Diagram shows viral life cycle from viral attachment and entry into host cells (1, 2), genome release from the capsid (3, 4), genome replication, transcription and protein expression (5, 6), and viral assembly and release (7, 8). (B) ASO-based antiviral strategies. Examples are given for four different types of viruses. Coronavirus (positive-strand RNA virus): ASOs target the transcription regulatory sequence (TRS) of the RNA genome (+RNA) to reduce viral subgenomic RNA production. Influenza (negative-strand RNA virus): ASOs target viral mRNAs to reduce the production of viral nucleoprotein and matrix protein. HBV (partially double-stranded DNA virus): ASOs target a conserved sequence of viral mRNAs to reduce the translation of viral proteins. HIV (retrovirus): ASOs bind to the viral genome to interfere with reverse transcription and hence reduce viral DNA production. Abbreviations: L, leader sequence; cRNA, complementary RNA; rcDNA, relaxed circular DNA; cccDNA, covalently closed circular DNA; pgRNA, pregenomic RNA.
2.1. Coronaviruses
The COVID-19 pandemic has prompted the rapid development of therapeutic strategies against SARS-CoV-2. A combination of cryo-electron microcopy and molecular modeling has revealed the tertiary structure of the frameshift stimulation element of SARS-CoV-2. LNA-modified ASOs targeting the structure of this element can disrupt translational frameshifting and hence inhibit viral replication [38][64]. Another report showed that a 2′-OMe/SP-ASO conjugated with four 2′-5′-oligoadenylates that can induce RNase L-mediated cleavage and degradation of the SARS-CoV-2 envelop and spike RNAs can effectively inhibit viral propagation in pseudovirus infection models [39][65].
2.2. Dengue Virus
Dengue infection occurs in tropical and subtropical areas and causes fever and flu-like symptoms. Dengue viruses have a 10.7 kb positive-strand RNA genome encoding three structural proteins and seven nonstructural proteins. The 5′ and 3′ untranslated regions, respectively, fold into conserved structures that are essential for viral viability. The 5′-most stem-loop acts as a promoter of viral RNA replication. RNA replication involves genome cyclization, which is mediated by the interaction between the complementary 5′ and 3′ cyclization sequences [40][66]. Arginine-rich CPP-PMOs that respectively target the 5′ or 3′-terminal stem-loop or 3′ cyclization sequence of the Dengue genome can inhibit viral replication and decrease viral titer in cultured cells [41][42][67,68]. Analogously, a recent study showed that 3′ stem-loop-targeting vivo-PMOs can potently inhibit Dengue replication in dendritic cells that are primary target cells of dengue infection [43][69].
2.3. Respiratory Syncytial Virus (RSV)
RSV causes lower respiratory tract disease that most often affects children and older individuals. RSV has a negative-strand RNA genome of ~15 kb. After entry into the host cell, the viral nucleocapsid and polymerase are delivered into the cytoplasm. Viral RNA-dependent RNA polymerase (RdRp) transcribes the viral genome into mRNAs that encode viral proteins and synthesizes the antigenome, which serves as the template for genome synthesis [44][70]. ASOs that induce RNase H-mediated cleavage/degradation of RSV genomic RNA can inhibit RSV replication [45][71]. A CPP-conjugated PS-PMO could inhibit RSV replication in mice by suppressing the translation of RSV L mRNA [46][72]. The intranasal route is a rational choice for delivery of antiviral drugs against respiratory infections. One study has demonstrated that intranasal administration of siRNAs that knock down RSV phosphoprotein expression can effectively reduce RSV infection and prevent pulmonary pathology in mice [47][73].
2.4. Influenza
Influenza viruses cause a contagious respiratory illness ranging from mild to severe. The influenza genome comprises eight negative-strand RNA segments, each of which encodes one or two proteins. In contrast to most RNA viruses, the influenza RNAs are transcribed and replicated by viral RdRp in the nucleus. For viral synthesis, RdRp uses a cap-snatching mechanism to prime transcription [48][74]. Notably, all eight viral RNAs contain conserved sequences respectively at their 5’ and 3’ termini [48][74]. Although neuraminidase inhibitors are the most frequently used anti-influenza drugs, other antiviral strategies are still necessary [49][75]. CPP-conjugated PMOs targeting the 3′ conserved region of the nucleocapsid mRNA can reduce the viral titer [50][76]. Using titanium dioxide (TiO2) as a nanocarrier, polylysine-linked ASOs targeting the same conserved region exhibited potent antiviral activity with little cytotoxicity [51][77].
2.5. Ebola Virus
3.5. Ebola Virus
Ebola virus is a rare but deadly virus that causes coagulation abnormalities, leading to hemorrhagic fever. A recent outbreak occurred in West Africa from 2014 to 2016. Ebola and its relative of the Marburg virus belong to the Filoviridae family, and these viruses have a negative-strand RNA genome of 19 kb encoding seven proteins. Among them, VP24 and VP35 antagonize the innate antiviral immune response via multiple pathways and are responsible for the extreme virulence of Ebola virus [52][80]. Essentially, VP24 inhibits the activation of interferon-stimulated genes by preventing nuclear import of a key transcription factor STAT1, whereas VP35 interacts with double-stranded RNA ends to prevent sensing by cellular pattern recognition receptors such as retinoic acid inducible gene-I (RIG-I) [53][54][81,82]. Positively charged PMOs targeting VP35 mRNA could protect mice from infection-induced lethality [55][83]. Moreover, targeting both VP24 and VP35 achieved postexposure efficacy against Ebola virus in nonhuman primates, indicating positively charged PMOs as effective therapeutic agents [56][84].
2.6. HBV
3.6. HBV
HBV, a prototype virus of the Hepadnaviridae family, has a 3.2 kb partially double-stranded, relaxed circular DNA genome [57][85]. After infection, the viral genome is converted to covalently closed circular DNA in the nucleus, and this DNA serves as the template for synthesis of pregenomic RNA and subgenomic viral transcripts. Viral replication occurs by reverse transcription of pregenomic RNA. Chronic HBV infection may lead to liver failure and liver cancer. Interferon and nucleoside/nucleotide analogs are the most commonly used therapeutics [58][86]. Developing new therapeutic strategies is still necessary, however, owing to drug resistance. In a pioneering study, a PS-ASO complementary to the HBV polyadenylation signal sequence complexed to a ASGPR ligand and polylysine reduced viral surface antigen (HBsAg) expression and blocked HBV replication in cultured cells [59][87]. Recently, GalNAc-conjugated LNA-ASOs that can destroy viral mRNAs via RNase-H-mediated degradation showed a significant HBsAg reduction in HBV-infected mice [60][88]. Notably, the uncapped 5′-triphosphate of RNA can activate the RIG-I-dependent antiviral type-I interferon response [61][89], and a recent study demonstrated that 5′-triphosphate-modified siRNAs can target HBV and meanwhile elicit an antiviral immune response [62][90].
2.7. HIV
3.7. HIV
HIV causes life-threatening immunodeficiency syndrome and thus has long attracted innovative development of antiviral drugs. HIV is a lentivirus that infects and subsequently depletes CD4+ helper T cells. Upon entry into these T cells, the viral RNA genome is reverse transcribed into DNA by viral reverse transcriptase. The resulting viral double-stranded DNA is integrated into the host genome by the viral integrase and host factors [63][92]. Many anti-HIV drugs have been developed to target viral enzymes (such as reverse transcriptase, integrase and protease) or prevent viral entry by blocking the T-cell receptors CD4 or CCR5 [64][93]. Numerous nucleic acid-based drugs have also been designed for HIV treatment. For example, early studies showed that an siRNA targeting the HIV genome or HIV Gag-p24 could inhibit viral replication [65][66][94,95].
3. ASOs Targeting Host Factors
3.1. Niemann-Pick C1 (NPC1)
NPC1 is a multi-transmembrane protein essential for cholesterol transport from late endosomes and lysosomes and regulates cellular lipid homeostasis [67][98]. NPC1 mutations cause accumulation of cholesterol and other lipids in various tissues. The lysosomal accumulation of lipids in Niemann-Pick type C disease with NPC1 mutations leads to neurological impairments and progressive neurodegeneration [68][99]. Moreover, NPC1 and NPC1-like protein participate in the infection by various viruses. NPC1 serves as a fusion receptor for filovirus. Ebola virus is internalized via a micropinocytosis-like process and is subsequently transported to late endosomes [69][100]. The Ebola virus glycoprotein (GP) is required for virion/cellular membrane fusion. Upon proteolytic processing of the GP1 subunit, its receptor-binding domain interacts with endosomal NPC1 for viral entry and subsequent release of the viral nucleoprotein in the cytoplasm followed by replication of viral genomic RNA [69][100]. Fibroblasts derived from patients with Niemann-Pick type C disease are resistant to filovirus infection [70][101]. LNA-PS-modified ASOs targeting NPC1 mRNA can interfere with the cellular entry of a filovirus glycoprotein-pseudotyped virus [71][102].
3.2. Raf-1
4.2. Raf-1
The serine/threonine kinase Raf1 is a downstream effector of RAS in the mitogen-activated protein kinase pathway. Ras/Raf/MEK/ERK signaling regulates numerous cellular processes such as proliferation and differentiation in response to extracellular stimuli [72][106]. A number of viruses exploit this pathway to modulate their infectious cycle. HCV activates Raf-1 by its core protein and thereby regulates hepatocyte growth and differentiation [73][107]. Conversely, Raf-1 participates in HCV replication via its interaction with the viral nonstructural protein 5A in the replication complex [74][108]. Thus, inhibition of Raf-1 attenuates viral replication. Notably, activation of the Ras/Raf/MEK pathway downregulates the expression of interferon-stimulated genes that are critical for the innate immune response. Therefore, HCV infection attenuates interferon signaling by activating Raf-1 and hence benefits viral propagation [75][109].3.3. The Heat-Shock Protein MRJ
4.3. The Heat-Shock Protein MRJ
Heat-shock proteins function as molecular chaperones to maintain proteostasis [76][113]. A number of viruses modulate the cellular heat-shock response or take advantage of cellular heat-shock proteins to overcome host environmental challenges and complete their life cycle [77][78][114,115]. An early study revealed that the human heat-shock protein DNAJB6 (also termed MRJ) is critical for nuclear import of the HIV-2 preintegration complex via its interaction with viral Vpx protein [79][116]. Notably, MRJ has two splice isoforms that exert different effects on viral infection [78][115]. Isoform switching from the C-terminally truncated MRJ-S to full-length MRJ-L occurs during monocyte differentiation into macrophages, which are target cells for HIV [80][117]. Accordingly, individuals with a higher level of MRJ-L in macrophages are more susceptible to HIV infection [81][118]. MRJ-L possibly facilitates the nuclear import of the HIV preintegration complex via its C-terminal nuclear localization signal [81][118]. Similarly, it promotes nuclear entry of the cytomegalovirus primase [82][119]. MRJ-L is also essential for subgenomic mRNA production and viral propagation of RSV even though the RSV life cycle is completed in the cytoplasm [80][117].3.4. miR-122
4.4. miR-122
miRNAs regulate gene expression at the post-transcriptional level via binding to the 3′ untranslated region of target mRNAs. Therefore, miRNAs can regulate the pathogenesis of a broad range of viruses; most of them downregulate viral translation and replication [83][121]. However, viruses can modulate the expression of host miRNAs, most of which are involved in antiviral innate immunity. Notably, HCV infection upregulates miR-122, which is abundant in hepatocytes and regulates liver homeostasis [84][122]. miR-122 in turn binds to sites upstream of the internal ribosome entry site in the 5′ non-coding region of the viral RNA genome and hence increases viral RNA stability and upregulates viral RNA translation and replication [85][86][123,124]. Host factors that are involved in miRNA biogenesis, such as Dicer, TRBP and Ago2, also contribute to viral RNA accumulation [87][125].3.5. Other Host Factors Targeted by ASOs
4.5. Other Host Factors Targeted by ASOs
ASGPR is a candidate receptor for HBV, and its major subunit ASGPR1 is upregulated in cells of HBV-infected patients. ASOs targeting ASGPR1 mRNA in HBV-infected human hepatocellular carcinoma cells can reduce the level of viral antigens and DNA [88][131]. A subcellular proteomic screen implicated the involvement of programmed cell death 5 (PDCD5) during influenza virus infection. PDCD5 may suppress tumors and pathogenic T cells by inhibiting cell proliferation and inducing apoptosis. Knockdown of PDCD5 mitigated influenza HIN1 propagation in cultured cells [89][132]. A subsequent study showed that PDCD5-targeting ASOs can downregulate PDCD5 in lung tissue and hence protect mice from influenza virus infection [90][133].