Compartmentalization of the membrane is essential for cells to perform highly specific tasks and spatially constrained biochemical functions in topographically defined areas. These membrane lateral heterogeneities range from nanoscopic dimensions, often involving only a few molecular constituents, to micron-sized mesoscopic domains resulting from the coalescence of nanodomains. Short-lived domains lasting for a few milliseconds coexist with more stable platforms lasting from minutes to days. This panoply of lateral domains subserves the great variety of demands of cell physiology, particularly high for those implicated in signaling. The dendritic spine, a subcellular structure of neurons at the receiving (postsynaptic) end of central nervous system excitatory synapses, exploits this compartmentalization principle. In its most frequent adult morphology, the mushroom-shaped spine harbors neurotransmitter receptors, enzymes, and scaffolding proteins tightly packed in a volume of a few femtoliters. In addition to constituting a mesoscopic lateral heterogeneity of the dendritic arborization, the dendritic spine postsynaptic membrane is further compartmentalized into spatially delimited nanodomains that execute separate functions in the synapse.
- dendritic spine
- plasma membrane
- membrane domains
1. Introduction
2. Dendritic Spines, Discrete Subcellular Compartment of the Neuronal Membrane
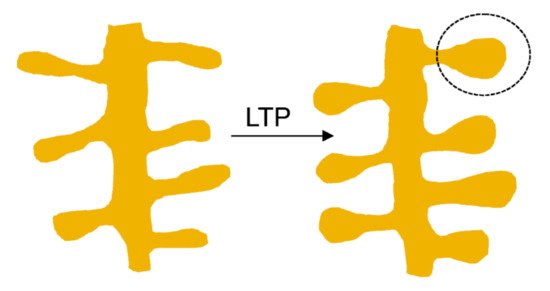
Dendritic spines constitute fundamental units of information endowed with processing synaptic (mostly excitatory) chemical neurotransmission in the mammalian brain. Spines compartmentalize biochemical and electrical signals, thus modulating the functional properties of synapses. Based on the relative length of their neck and the diameter of their head, dendritic spines have been arbitrarily classified into five categories: mushroom, thin, stubby, filopodia, and bifurcation- or cup-shaped spines [15][9]. However, this represents a fragmentary, anatomically static view of the dendritic spine: in fact, they also have a dynamic dimension, allowing them to modify their size and shape within seconds to minutes and undergo more lasting changes in the time scale of hours to days. In mature neurons spine motility declines [13][7]. Spine head volumes range from 0.01 to 1 μm3, and spine necks vary between 50 and 500 nm in diameter and up to 3 μm in length [16,17,18][10][11][12]. The size of the spine head has been shown to correlate with the size of the postsynaptic density (PSD) a characteristic morphological differentiation of the postsynapse [16[10][11][12],17,18], and with the amplitude of the excitatory postsynaptic current (EPSC) [19,20][13][14]. One of the most important functions of the brain is enabling the neural activity generated by an experience to modify neural circuit functionality in a process referred to as synaptic plasticity. This property of individual synapses affects higher brain functions like thought, feeling, and behavior [21][15]. Morphological correlates of synaptic plasticity at the dendritic spine level have been described, involving the remodeling of the PSD to regulate the number of intervening receptors and scaffolding proteins. The so-called long-term potentiation (LTP) and long-term depression (LTD), two mechanisms that have been thoroughly studied in the mammalian hippocampus, are induced by recent patterns of activity that strengthen or depress synapse activity, respectively [22][16] (Figure 1).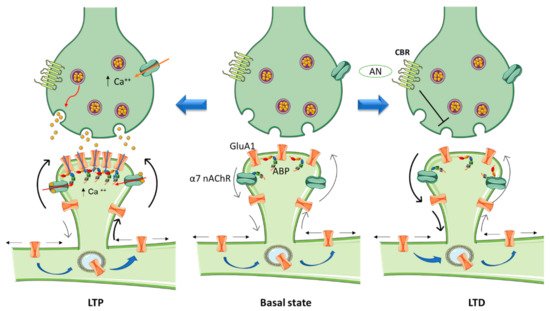
3. Cannabinoids and Cannabinoid Receptors
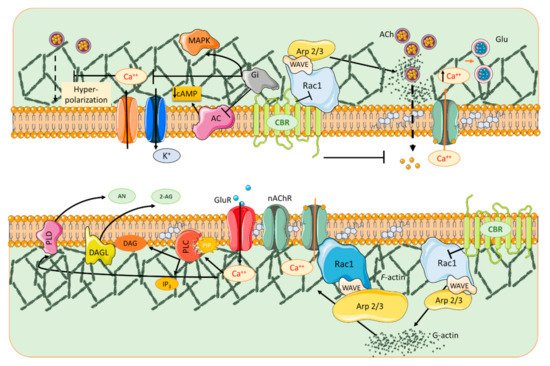
Cannabinoids have been shown to alter the organization of the actin cytoskeleton in various cell types [58][27]. Njoo and collaborators highlighted the functional significance of CBR1 interactions with the Wiskott-Aldrich syndrome protein-family verprolin-homologous protein 1 (WAVE1)/SCAR1 complex [59][28]. These authors stated that the complex plays a key role in dynamically regulating the actin cytoskeleton in developing and adult neurons, thus contributing to the most salient functions of eCBs in the brain and spinal cord. Furthermore, they showed that cannabinoids structurally remodel dendritic spines by regulating the activity levels of WAVE1, and reported a novel function for WAVE1 in mediating inflammatory pain via structural and functional plasticity of spinal neurons [59][28]. Although CBR1s are mainly expressed presynaptically, several studies have indicated that CBR1 can be additionally localized in neuronal dendrites [60,61][29][30] colocalizing with the PSD protein PSD-95 in spines [59][28]. Rac1 activity decreases within minutes of CBR1 activation, limiting the conversion of G-actin to F-actin in dendritic spines of mature cortical neurons. This in turn produces a depletion of mature spines with the characteristic adult-type, elaborate mushroom morphology, which are believed to mediate increased synaptic efficacy such as observed in LTP [62][31]. In agreement with these findings, chronic treatment with the synthetic cannabinoid agonist WIN 55212-2 (WIN) reduced spine density in the nucleus accumbens (Nac) 24 h after the last injection [63][32]. Spiga and collaborators [64,65][33][34] also reported a decrease in spine density in the Nac (core) after a 1 h withdrawal period. Activation of CBR1 in hippocampal neurons elicits a decrease in presynaptic F-actin and other cytoskeletal proteins, including ARPC2 and WASF1/WAVE1 that correlate with morphological changes (e.g., reduction in bouton size) [66][35], as seen at the postsynapse.4. Cholinergic Signaling Contribution to Glutamatergic Receptor Compartmentalization
Crosstalk between the dopaminergic and glutamatergic systems enables them to initiate and organize normal behavior [90][36]. NMDARs can act as a scaffold to recruit laterally diffusing dopamine D1 receptors (D1Rs) to spines. The activation of these NMDARs alters the topography and movement of D1Rs by trapping them in dendritic spines [91][37]. D1Rs selectively interact with the NR1 subunit of the NMDAR through its C-terminal tail to form dimeric hetero-complexes. Induction of LTP in the striatum requires activation of D1Rs since antagonizing these receptors blocks NMDAR–dependent LTP; while in the cortex, working memory is altered by this antagonism [92,93,94][38][39][40]. Activation of D1Rs via DA release, caused e.g., by cholinergic signaling activation, can then recruit D1R-NMDAR complexes in a regulated manner [95][41]. Cholinergic signaling pathways thus contribute to sub-compartmentalize glutamatergic neurotransmission, spatially restricting its sphere of action. More generally, the ubiquitous distribution of nicotinic receptors in brain enables them to regulate many important high-level cognitive functions such as attention, working memory, learning processes [96][42], cognitive flexibility [97][43] and social interactions [98][44]. They are also involved in addiction and dependence [81,82][45][46].65. Cannabinoids and Nicotinic Receptors
In the activated spine, Ca2+ entry through postsynaptic α7 nAChRs [70][47] or NMDA receptors and/or voltage-gated Ca2+ channels [124][48] can promote the activation of multiple signaling pathways with specific spatio-temporal patterns that orchestrate and regulate different aspects of cytoskeletal dynamics in the stimulated spines. Within the spine, Ca2+ binds to calmodulin (CaM), a Ca2+-binding protein which subsequently activates Ca2+/CaM-dependent kinases and phosphatases such as CaMKII and calcineurin [125,126][49][50]. CaMKII activates small GTPases and these in turn further modulate several downstream kinases [127][51] that have the ability to activate many ABPs, including Cofilin and Arp2/3, two proteins that play essential roles in actin remodeling [128,129][52][53]. Thus cannabinoids, through rac1/WAVE 1 modulation and α7 nAChR activation, contribute to actin structural remodeling of dendritic spines. Whereas activation of Rac1/WAVE1 induces a depletion of mushroom type spines, α7 nAChR activation contributes to the formation and maturation of dendritic spines (Figure 1).
76. Importance of Lipids in Dendritic Spine Compartmentalization
At the synapse, cholesterol interacts with several neurotransmitter receptors through consensus linear binding sequences like the so-called cholesterol recognition/interaction amino acid consensus motifs (CRAC and its mirror image CARC [143][54]. These consensus domains have been proposed to facilitate membrane protein incorporation into cholesterol-rich domains in a great variety of membrane proteins, including the superfamily of pentameric ligand-gated ion channels (pLGICs) and the superfamily of G-protein coupled receptors (GPCRs). The prototypic LGIC, the nAChR, exhibits a CRAC motif adjacent to the transmembrane helix M1, and a CARC sequence on the M4-facing surface of M1 adjacent to one of the proposed cholesterol-binding cavities [143,144][54][55]. Likewise, the transmembrane helix 7 of human cannabinoid receptor 1 (CBR1) displays a CRAC sequence [145][56]. A cholesterol molecule was recently identified in a crystal structure [146][57] and a cryo-electron microscopy (EM) structure [147][58] of the CBR1. Sphingolipids participate as plasma membrane lipids and signaling molecules such as ceramide, sphingosine, and sphingosine-1-phosphate that are produced after the metabolism of plasma membrane sphingolipids [148][59]. Among sphingolipids, sphingomyelins are enriched in brain membranes. The elimination of dendritic spines upon reduction of cholesterol and sphingomyelin levels was described almost 20 years ago [149][60], highlighting the importance of these lipids for neuronal communication. Ceramide promotes spine maturation by contributing to the transformation of dendritic filopodia to mature spines [150][61]. Phosphoinositides (PIPs) are important players in postsynaptic excitability since they have an exceptional high rate of metabolic turnover and compartmentalization [151][62]. There are multiple enzymes at dendritic spines that interconvert different PIPs contributing to the dynamic role of lipids in the plasma membrane. Phosphatidylinositol (4,5) diphosphate (PIP2) is converted to phosphatidylinositol (3,4,5) triphosphate (PIP3) by phosphatidylinositol-4,5-bisphosphate 3-kinase. PIP3 content at the spines is higher than that in dendritic shafts under basal conditions. Furthermore, upon glutamate stimulation PIP3 redistributes contributing to the formation of fine spinules projecting from spines [152,153][63][64]. Additionally, PIP3, because of its capacity regulate the activity of multiple Rho GTPase effectors [154][65], has also been implicated in the interaction of membrane-cytoskeleton crosstalk at spines, and is able to regulate the Akt-mTOR pathway to participate in dendritic spine morphogenesis [155,156][66][67]. The PIP2-clustering molecule myristoylated alanine-rich C kinase substrate (MARCKS) reversibly sequesters PIP2 on the plasma membrane, upon local increases in intracellular calcium [157][68]. MARCKs contributes to spine morphogenesis by promoting the transition from immature dendritic spines to larger and more stable mushroom-shaped spines by controlling actin cytoskeleton [158][69]. Moreover, association of MARCKS to cholesterol at the membrane is necessary for its ability to crosslink F-actin [158][69]. In addition, PIP3 contributes to the accumulation of PSD-95 at spines whereas conversion of PIP2 by phospholipase C favors synaptic actin depolymerization and PSD-95 degradation, thus contributing to spine remodeling [159][70]. Among other factors, actin dynamics is modulated by membrane lipids. A reduction in membrane cholesterol levels produces a rapid collapse of spine morphological integrity associated with redistribution of F-actin from the spine proper to the dendritic shaft [149][60]. Sphingolipids also play a relevant role in the spine plasma membrane-actin cytoskeleton crosstalk. Sphingomyelins modulate membrane binding and activity of the Rho GTPases, key regulators of the actin cytoskeleton in the synapse. Accumulation of sphingomyelins at postsynaptic membranes, as observed in a Niemann-Pick disease type A mouse model defective in acid sphingomyelinase, induces a reduction of metabotropic glutamate receptors that impairs the membrane attachment of RhoA and its effectors ROCK (RhoA kinase) and profilin IIa. This impairment results in the diminution of F-actin content, ultimately reducing spine number and size [160][71]. The conversion of sphingomyelin to ceramide at the plasma membrane is catalyzed by neutral sphingomyelinase-2 [161][72]. This enzyme can, in turn, modulate spine actin cytoskeleton. Activation of the neutral sphingomyelinase restores F-actin content of dendritic spines by enhancing the RhoA pathway in mice defective of the acid sphingomyelinase, which present high sphingomyelin synaptic levels [160][71]. Conversely, inhibition of the sphingomyelinase decreases the abnormally high levels of F-actin in spines of neurons in mice lacking Wiskott–Aldrich syndrome protein interacting protein (WIP) [162][73]. Thus, through various protein and lipid interactions, actin promotes compartmentalization of the plasma membrane and has an important role in the modulation of neuronal strength.Impact of the Lipid Microenvironment on nAChRs and CBRs
The best documented example of the influence that the lipid microenvironment and cholesterol exert on the topography and function of a neurotransmitter receptor is provided by the paradigm rapid LGIC, the nAChR (see reviews in [163,164][74][75]). Changes in cholesterol levels alter the translational mobility of the receptor in the plane of the plasma membrane, as measured by fluorescence recovery after photobleaching and fluorescence correlation spectroscopy [165][76] and single-molecule localization microscopy [166,167][77][78]. Cell-surface trafficking of nAChRs is dependent on cholesterol metabolism [168,169][79][80]. Pharmacological long-term inhibition of cholesterol biosynthesis by the statin lovastatin differentially augments cell-surface levels of α4β2 and α7 nAChRs in neurites and soma of rat hippocampal neurons [169][80]. Misbalances in brain cholesterol homeostasis affect cholinergic signaling involving α4β2 and α7 nAChRs and indirectly impact on the number and distribution of other neuroreceptors at dendritic spines, with important consequences for brain function. Sphingolipids are also necessary for nAChR export in the early secretory pathways [170][81]. Thus, any modification in sphingolipid levels will impact on nAChR expression.87. Contribution of Actin Dynamics to the Compartmentalization of the Dendritic Spine
98. Nanodomain Cluster Organization Emerges as a Common Organizing Principle at CNS Synapses
Neuronal activity regulates receptor fluxes at the PSD through the interaction of receptors with scaffolding molecules and lipids [193][101]. The PSD scaffolding proteins are distributed heterogeneously and form nanodomains within the synapses, termed sub-synaptic domains (SSD) [194][102]. The characteristic output of LTP induction, spine enlargement, is paralleled by the increase in the number of PSD-95 (an SSD-resident protein) copies [195][103]. This augmentation in PSD-95 biosynthesis occurs within hours of LTP induction [196][104]. During development, proteins organized in nanoclusters at the pre- and postsynapse control the differentiation of the dendritic spine. The cholesterol-binding protein TSPAN5, which belongs to the tetraspanin superfamily, is localized postsynaptically in pyramidal excitatory neurons, and is the master controller of spine maturation. Control is exerted by promoting the clustering of the postsynaptic molecule neuroligin-1 at the spine surface [197][105], which in turn recognizes and binds to presynaptic neurexins. TSPAN5 knockout mice have an extremely low number of spines, reinforcing the view that this tetraspanin plays a key role in excitatory synapse maturation. At excitatory glutamatergic synapses, the lateral exchange of receptors between extrasynaptic and synaptic areas is responsible for neuronal plasticity [193,201,202][101][106][107]. In the CNS, endocytosis occurs predominantly, if not exclusively, in extrasynaptic areas [203[108][109][110],204,205], and hence receptors have to diffuse out of the PSD to be internalized [206][111]. In turn, mobilization of receptors from the spine shaft to the PSD partly depends on the morphology of the spine. As the spine neck diameter diminishes, it is energetically and mechanically more costly to transport endosomal vesicles from the ER outposts in the dendrite to the PSD through the spine actin meshwork [207][112]. Dendritic ER and Golgi outposts are well documented in rodent hippocampal neurons, particularly at dendritic branching points [208][113] and high-pressure freezing fixation (without chemical fixation) combined with transmission electron microscopy has revealed smooth ER and Golgi outposts inside dendritic spines [209][114] (Figure 4).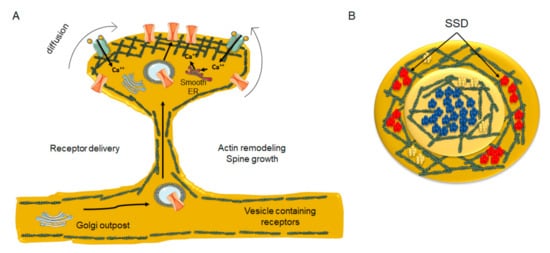
109. Concluding Remarks
References
- Brusés, J.L.; Chauvet, N.; Rutishauser, U. Membrane lipid rafts are necessary for the maintenance of the (alpha)7 nicotinic acetylcholine receptor in somatic spines of ciliary neurons. J. Neurosci. 2001, 21, 504–512.
- Hayashi, T.; Su, T.-P. Sigma-1 receptors at galactosylceramide-enriched lipid microdomains regulate oligodendrocyte differentiation. Proc. Natl. Acad. Sci. USA 2004, 101, 14949–14954.
- Allen, J.A.; Halverson-Tamboli, R.A.; Rasenick, M.M. Lipid raft microdomains and neurotransmitter signalling. Nat. Rev. Neurosci. 2007, 8, 128–140.
- Egawa, J.; Pearn, M.L.; Lemkuil, B.P.; Patel, P.M.; Head, B.P. Membrane lipid rafts and neurobiology: Age-related changes in membrane lipids and loss of neuronal function. J. Physiol. 2016, 594, 4565–4579.
- Nagappan, G.; Lu, B. Activity-dependent modulation of the BDNF receptor TrkB: Mechanisms and implications. Trends Neurosci. 2005, 28, 464–471.
- Fernandes, C.C.; Berg, D.K.; Gómez-Varela, D. Lateral mobility of nicotinic acetylcholine receptors on neurons is determined by receptor composition, local domain, and cell type. J. Neurosci. 2010, 30, 8841–8851.
- Dunaevsky, A.; Tashiro, A.; Majewska, A.; Mason, C.; Yuste, R. Developmental regulation of spine motility in the mammalian central nervous system. Proc. Natl. Acad. Sci. USA 1999, 96, 13438–13443.
- Heine, M.; Holcman, D. Asymmetry between Pre- and Postsynaptic Transient Nanodomains Shapes Neuronal Communication. Trends Neurosci. 2020, 43, 182–196.
- Hering, H.; Sheng, M. Dentritic spines: Structure, dynamics and regulation. Nat. Rev. Neurosci. 2001, 2, 880–888.
- Trommald, M.; Hulleberg, G. Dimensions and density of dendritic spines from rat dentate granule cells based on reconstructions from serial electron micrographs. J. Comp. Neurol. 1997, 377, 15–28.
- Arellano, J.I.; Benavides-Piccione, R.; Defelipe, J.; Yuste, R. Ultrastructure of dendritic spines: Correlation between synaptic and spine morphologies. Front. Neurosci. 2007, 1, 131–143.
- Harris, K.M.; Stevens, J.K. Dendritic spines of CA 1 pyramidal cells in the rat hippocampus: Serial electron microscopy with reference to their biophysical characteristics. J. Neurosci. 1989, 9, 2982–2997.
- Noguchi, J.; Nagaoka, A.; Watanabe, S.; Ellis-Davies, G.C.R.; Kitamura, K.; Kano, M.; Matsuzaki, M.; Kasai, H. In vivo two-photon uncaging of glutamate revealing the structure-function relationships of dendritic spines in the neocortex of adult mice. J. Physiol. 2011, 589, 2447–2457.
- Matsuzaki, M.; Ellis-Davies, G.C.R.; Nemoto, T.; Miyashita, Y.; Iino, M.; Kasai, H. Dendritic spine geometry is critical for AMPA receptor expression in hippocampal CA1 pyramidal neurons. Nat. Neurosci. 2001, 4, 1086–1092.
- Citri, A.; Malenka, R.C. Synaptic Plasticity: Multiple Forms, Functions, and Mechanisms. Neuropsychopharmacology 2008, 33, 18–41.
- Bliss, T.V.P.; Cooke, S.F. Long-term potentiation and long-term depression: A clinical perspective. Clinics 2011, 66 (Suppl. 1), 3–17.
- Matsuzaki, M.; Honkura, N.; Ellis-Davies, G.C.R.; Kasai, H. Structural basis of long-term potentiation in single dendritic spines. Nature 2004, 429, 761–766.
- Lang, C.; Barco, A.; Zablow, L.; Kandel, E.R.; Siegelbaum, S.A.; Zakharenko, S.S. Transient expansion of synaptically connected dendritic spines upon induction of hippocampal long-term potentiation. Proc. Natl. Acad. Sci. USA 2004, 101, 16665–16670.
- Zhou, Q.; Homma, K.J.; Poo, M. Shrinkage of dendritic spines associated with long-term depression of hippocampal synapses. Neuron 2004, 44, 749–757.
- Nägerl, U.V.; Eberhorn, N.; Cambridge, S.B.; Bonhoeffer, T. Bidirectional activity-dependent morphological plasticity in hippocampal neurons. Neuron 2004, 44, 759–767.
- Tønnesen, J.; Katona, G.; Rózsa, B.; Nägerl, U.V. Spine neck plasticity regulates compartmentalization of synapses. Nat. Neurosci. 2014, 17, 678–685.
- Vanderklish, P.W.; Edelman, G.M. Dendritic spines elongate after stimulation of group 1 metabotropic glutamate receptors in cultured hippocampal neurons. Proc. Natl. Acad. Sci. USA 2002, 99, 1639–1644.
- Tønnesen, J.; Nägerl, U.V. Dendritic Spines as Tunable Regulators of Synaptic Signals. Front. Psychiatry 2016, 7, 101.
- Müller, W.; Connor, J.A. Dendritic spines as individual neuronal compartments for synaptic Ca2+ responses. Nature 1991, 354, 73–76.
- Guthrie, P.B.; Segal, M.; Kater, S.B. Independent regulation of calcium revealed by imaging dendritic spines. Nature 1991, 354, 76–80.
- Honigmann, A.; Pralle, A. Compartmentalization of the Cell Membrane. J. Mol. Biol. 2016, 428, 4739–4748.
- Hohmann, T.; Feese, K.; Ghadban, C.; Dehghani, F.; Grabiec, U. On the influence of cannabinoids on cell morphology and motility of glioblastoma cells. PLoS ONE 2019, 14, e0212037.
- Njoo, C.; Agarwal, N.; Lutz, B.; Kuner, R. The Cannabinoid Receptor CB1 Interacts with the WAVE1 Complex and Plays a Role in Actin Dynamics and Structural Plasticity in Neurons. PLOS Biol. 2015, 13, e1002286.
- Ladarre, D.; Roland, A.B.; Biedzinski, S.; Ricobaraza, A.; Lenkei, Z. Polarized cellular patterns of endocannabinoid production and detection shape cannabinoid signaling in neurons. Front. Cell. Neurosci. 2014, 8, 426.
- Leterrier, C.; Lainé, J.; Darmon, M.; Boudin, H.; Rossier, J.; Lenkei, Z. Constitutive activation drives compartment-selective endocytosis and axonal targeting of type 1 cannabinoid receptors. J. Neurosci. 2006, 26, 3141–3153.
- Nimchinsky, E.A.; Sabatini, B.L.; Svoboda, K. Structure and function of dendritic spines. Annu. Rev. Physiol. 2002, 64, 313–353.
- Carvalho, A.F.; Reyes, B.A.S.; Ramalhosa, F.; Sousa, N.; Van Bockstaele, E.J. Repeated administration of a synthetic cannabinoid receptor agonist differentially affects cortical and accumbal neuronal morphology in adolescent and adult rats. Brain Struct. Funct. 2016, 221, 407–419.
- Spiga, S.; Lintas, A.; Diana, M. Altered Mesolimbic Dopamine System in THC Dependence. Curr. Neuropharmacol. 2011, 9, 200–204.
- Spiga, S.; Lintas, A.; Migliore, M.; Diana, M. Altered architecture and functional consequences of the mesolimbic dopamine system in cannabis dependence. Addict. Biol. 2010, 15, 266–276.
- Monday, H.R.; Bourdenx, M.; Jordan, B.A.; Castillo, P.E. Cb1-receptor-mediated inhibitory ltd triggers presynaptic remodeling via protein synthesis and ubiquitination. Elife 2020, 9, 1–25.
- Scott, L.; Zelenin, S.; Malmersjö, S.; Kowalewski, J.M.; Markus, E.Z.; Nairn, A.C.; Greengard, P.; Brismar, H.; Aperia, A. Allosteric changes of the NMDA receptor trap diffusible dopamine 1 receptors in spines. Proc. Natl. Acad. Sci. USA 2006, 103, 762–767.
- Cepeda, C.; Levine, M.S. Where do you think you are going? The NMDA-D1 receptor trap. Sci. STKE 2006, 2006, pe20.
- Calabresi, P.; Gubellini, P.; Centonze, D.; Picconi, B.; Bernardi, G.; Chergui, K.; Svenningsson, P.; Fienberg, A.A.; Greengard, P. Dopamine and cAMP-regulated phosphoprotein 32 kDa controls both striatal long-term depression and long-term potentiation, opposing forms of synaptic plasticity. J. Neurosci. 2000, 20, 8443–8451.
- Kerr, J.N.; Wickens, J.R. Dopamine D-1/D-5 receptor activation is required for long-term potentiation in the rat neostriatum in vitro. J. Neurophysiol. 2001, 85, 117–124.
- Sawaguchi, T.; Goldman-Rakic, P.S. The role of D1-dopamine receptor in working memory: Local injections of dopamine antagonists into the prefrontal cortex of rhesus monkeys performing an oculomotor delayed-response task. J. Neurophysiol. 1994, 71, 515–528.
- Fiorentini, C.; Gardoni, F.; Spano, P.; Di Luca, M.; Missale, C. Regulation of dopamine D1 receptor trafficking and desensitization by oligomerization with glutamate N-methyl-D-aspartate receptors. J. Biol. Chem. 2003, 278, 20196–20202.
- Maurer, S.V.; Williams, C.L. The Cholinergic System Modulates Memory and Hippocampal Plasticity via Its Interactions with Non-Neuronal Cells. Front. Immunol. 2017, 8, 1489.
- Prado, V.F.; Janickova, H.; Al-Onaizi, M.A.; Prado, M.A.M. Cholinergic circuits in cognitive flexibility. Neuroscience 2017, 345, 130–141.
- Wang, L.; Almeida, L.E.F.; Spornick, N.A.; Kenyon, N.; Kamimura, S.; Khaibullina, A.; Nouraie, M.; Quezado, Z.M.N. Modulation of social deficits and repetitive behaviors in a mouse model of autism: The role of the nicotinic cholinergic system. Psychopharmacology 2015, 232, 4303–4316.
- Pidoplichko, V.I.; DeBiasi, M.; Williams, J.T.; Dani, J.A. Nicotine activates and desensitizes midbrain dopamine neurons. Nature 1997, 390, 401–404.
- Corrigall, W.A.; Franklin, K.B.; Coen, K.M.; Clarke, P.B. The mesolimbic dopaminergic system is implicated in the reinforcing effects of nicotine. Psychopharmacology 1992, 107, 285–289.
- Dani, J.A.; Bertrand, D. Nicotinic acetylcholine receptors and nicotinic cholinergic mechanisms of the central nervous system. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 699–729.
- Sabatini, B.L.; Oertner, T.G.; Svoboda, K. The life cycle of Ca(2+) ions in dendritic spines. Neuron 2002, 33, 439–452.
- Fujii, H.; Inoue, M.; Okuno, H.; Sano, Y.; Takemoto-Kimura, S.; Kitamura, K.; Kano, M.; Bito, H. Nonlinear decoding and asymmetric representation of neuronal input information by CaMKIIα and calcineurin. Cell Rep. 2013, 3, 978–987.
- Chang, J.-Y.; Parra-Bueno, P.; Laviv, T.; Szatmari, E.M.; Lee, S.-J.R.; Yasuda, R. CaMKII Autophosphorylation Is Necessary for Optimal Integration of Ca(2+) Signals during LTP Induction, but Not Maintenance. Neuron 2017, 94, 800–808.
- Murakoshi, H.; Wang, H.; Yasuda, R. Local, persistent activation of Rho GTPases during plasticity of single dendritic spines. Nature 2011, 472, 100.
- Borovac, J.; Bosch, M.; Okamoto, K. Regulation of actin dynamics during structural plasticity of dendritic spines: Signaling messengers and actin-binding proteins. Mol. Cell. Neurosci. 2018, 91, 122–130.
- Costa, J.F.; Dines, M.; Lamprecht, R. The Role of Rac GTPase in Dendritic Spine Morphogenesis and Memory. Front. Synaptic Neurosci. 2020, 12, 12.
- Baier, C.J.; Fantini, J.; Barrantes, F.J. Disclosure of cholesterol recognition motifs in transmembrane domains of the human nicotinic acetylcholine receptor. Sci. Rep. 2011, 1, 69.
- Fantini, J.; Di Scala, C.; Evans, L.S.; Williamson, P.T.F.; Barrantes, F.J. A mirror code for protein-cholesterol interactions in the two leaflets of biological membranes. Sci. Rep. 2016, 6, 21907.
- Oddi, S.; Dainese, E.; Fezza, F.; Lanuti, M.; Barcaroli, D.; De Laurenzi, V.; Centonze, D.; Maccarrone, M. Functional characterization of putative cholesterol binding sequence (CRAC) in human type-1 cannabinoid receptor. J. Neurochem. 2011, 116, 858–865.
- Hua, T.; Vemuri, K.; Pu, M.; Qu, L.; Han, G.W.; Wu, Y.; Zhao, S.; Shui, W.; Li, S.; Korde, A.; et al. Crystal Structure of the Human Cannabinoid Receptor CB(1). Cell 2016, 167, 750–762.
- Krishna Kumar, K.; Shalev-Benami, M.; Robertson, M.J.; Hu, H.; Banister, S.D.; Hollingsworth, S.A.; Latorraca, N.R.; Kato, H.E.; Hilger, D.; Maeda, S.; et al. Structure of a Signaling Cannabinoid Receptor 1-G Protein Complex. Cell 2019, 176, 448–458.
- Allende, M.L.; Zhu, H.; Kono, M.; Hoachlander-Hobby, L.E.; Huso, V.L.; Proia, R.L. Genetic defects in the sphingolipid degradation pathway and their effects on microglia in neurodegenerative disease. Cell. Signal. 2021, 78, 109879.
- Hering, H.; Lin, C.C.; Sheng, M. Lipid rafts in the maintenance of synapses, dendritic spines, and surface AMPA receptor stability. J. Neurosci. 2003, 23, 3262–3271.
- Carrasco, P.; Sahún, I.; McDonald, J.; Ramírez, S.; Jacas, J.; Gratacós, E.; Sierra, A.Y.; Serra, D.; Herrero, L.; Acker-Palmer, A.; et al. Ceramide levels regulated by carnitine palmitoyltransferase 1C control dendritic spine maturation and cognition. J. Biol. Chem. 2012, 287, 21224–21232.
- Hammond, G.R.V.; Schiavo, G. Polyphosphoinositol lipids: Under-PPInning synaptic function in health and disease. Dev. Neurobiol. 2007, 67, 1232–1247.
- Richards, D.A.; Mateos, J.M.; Hugel, S.; De Paola, V.; Caroni, P.; Gähwiler, B.H.; McKinney, R.A. Glutamate induces the rapid formation of spine head protrusions in hippocampal slice cultures. Proc. Natl. Acad. Sci. USA 2005, 102, 6166–6171.
- Ueda, Y.; Hayashi, Y. PIP3 regulates Spinule formation in dendritic spines during structural long-term potentiation. J. Neurosci. 2013, 33, 11040–11047.
- Yin, H.L.; Janmey, P.A. Phosphoinositide Regulation of the Actin Cytoskeleton. Annu. Rev. Physiol. 2003, 65, 761–789.
- Kumar, V.; Zhang, M.X.; Swank, M.W.; Kunz, J.; Wu, G.Y. Regulation of dendritic morphogenesis by Ras-PI3K-Akt-mTOR and Ras-MAPK signaling pathways. J. Neurosci. 2005, 25, 11288–11299.
- Kelleher, R.J., 3rd; Govindarajan, A.; Tonegawa, S. Translational regulatory mechanisms in persistent forms of synaptic plasticity. Neuron 2004, 44, 59–73.
- McLaughlin, S.; Murray, D. Plasma membrane phosphoinositide organization by protein electrostatics. Nature 2005, 438, 605–611.
- Calabrese, B.; Halpain, S. Essential role for the PKC target MARCKS in maintaining dendritic spine morphology. Neuron 2005, 48, 77–90.
- Horne, E.A.; Dell’Acqua, M.L. Phospholipase C is required for changes in postsynaptic structure and function associated with NMDA receptor-dependent long-term depression. J. Neurosci. 2007, 27, 3523–3534.
- Arroyo, A.I.; Camoletto, P.G.; Morando, L.; Sassoe-Pognetto, M.; Giustetto, M.; Van Veldhoven, P.P.; Schuchman, E.H.; Ledesma, M.D. Pharmacological reversion of sphingomyelin-induced dendritic spine anomalies in a Niemann Pick disease type A mouse model. EMBO Mol. Med. 2014, 6, 398–413.
- Stoffel, W. Functional analysis of acid and neutral sphingomyelinases in vitro and in vivo. Chem. Phys. Lipids 1999, 102, 107–121.
- Franco-Villanueva, A.; Fernández-López, E.; Gabandé-Rodríguez, E.; Bañón-Rodríguez, I.; Esteban, J.A.; Antón, I.M.; Ledesma, M.D. WIP modulates dendritic spine actin cytoskeleton by transcriptional control of lipid metabolic enzymes. Hum. Mol. Genet. 2014, 23, 4383–4395.
- Barrantes, F.J. Structural basis for lipid modulation of nicotinic acetylcholine receptor function. Brain Res. Brain Res. Rev. 2004, 47, 71–95.
- Barrantes, F.J. Cholesterol effects on nicotinic acetylcholine receptor. J. Neurochem. 2007, 103, 72–80.
- Baier, C.J.; Gallegos, C.E.; Levi, V.; Barrantes, F.J. Cholesterol modulation of nicotinic acetylcholine receptor surface mobility. Eur. Biophys. J. 2010, 39, 213–227.
- Mosqueira, A.; Camino, P.A.; Barrantes, F.J. Antibody-induced crosslinking and cholesterol-sensitive, anomalous diffusion of nicotinic acetylcholine receptors. J. Neurochem. 2020, 152, 663–674.
- Mosqueira, A.; Camino, P.A.; Barrantes, F.J. Cholesterol modulates acetylcholine receptor diffusion by tuning confinement sojourns and nanocluster stability. Sci. Rep. 2018, 8, 11974.
- Pediconi, M.F.; Gallegos, C.E.; De Los Santos, E.B.; Barrantes, F.J. Metabolic cholesterol depletion hinders cell-surface trafficking of the nicotinic acetylcholine receptor. Neuroscience 2004, 128, 239–249.
- Borroni, V.; Kamerbeek, C.; Pediconi, M.F.; Barrantes, F.J. Lovastatin Differentially Regulates α7 and α4 Neuronal Nicotinic Acetylcholine Receptor Levels in Rat Hippocampal Neurons. Molecules 2020, 25, 4838.
- Baier, C.J.; Barrantes, F.J. Sphingolipids are necessary for nicotinic acetylcholine receptor export in the early secretory pathway. J. Neurochem. 2007, 101, 1072–1084.
- Kapus, A.; Janmey, P. Plasma membrane--cortical cytoskeleton interactions: A cell biology approach with biophysical considerations. Compr. Physiol. 2013, 3, 1231–1281.
- Kusumi, A.; Nakada, C.; Ritchie, K.; Murase, K.; Suzuki, K.; Murakoshi, H.; Kasai, R.S.; Kondo, J.; Fujiwara, T. Paradigm shift of the plasma membrane concept from the two-dimensional continuum fluid to the partitioned fluid: High-speed single-molecule tracking of membrane molecules. Annu. Rev. Biophys. Biomol. Struct. 2005, 34, 351–378.
- Andrade, D.M.; Clausen, M.P.; Keller, J.; Mueller, V.; Wu, C.; Bear, J.E.; Hell, S.W.; Lagerholm, B.C.; Eggeling, C. Cortical actin networks induce spatio-temporal confinement of phospholipids in the plasma membrane--a minimally invasive investigation by STED-FCS. Sci. Rep. 2015, 5, 11454.
- Fujiwara, T.; Ritchie, K.; Murakoshi, H.; Jacobson, K.; Kusumi, A. Phospholipids undergo hop diffusion in compartmentalized cell membrane. J. Cell Biol. 2002, 157, 1071–1081.
- Auth, T.; Gov, N.S. Diffusion in a fluid membrane with a flexible cortical cytoskeleton. Biophys. J. 2009, 96, 818–830.
- Hotulainen, P.; Hoogenraad, C.C. Actin in dendritic spines: Connecting dynamics to function. J. Cell Biol. 2010, 189, 619–629.
- Kwik, J.; Boyle, S.; Fooksman, D.; Margolis, L.; Sheetz, M.P.; Edidin, M. Membrane cholesterol, lateral mobility, and the phosphatidylinositol 4,5-bisphosphate-dependent organization of cell actin. Proc. Natl. Acad. Sci. USA 2003, 100, 13964–13969.
- Cingolani, L.A.; Goda, Y. Actin in action: The interplay between the actin cytoskeleton and synaptic efficacy. Nat. Rev. Neurosci. 2008, 9, 344–356.
- Nelson, J.C.; Stavoe, A.K.H.; Colón-Ramos, D.A. The actin cytoskeleton in presynaptic assembly. Cell Adh. Migr. 2013, 7, 379–387.
- Michel, K.; Müller, J.A.; Oprişoreanu, A.-M.; Schoch, S. The presynaptic active zone: A dynamic scaffold that regulates synaptic efficacy. Exp. Cell Res. 2015, 335, 157–164.
- Rust, M.B.; Maritzen, T. Relevance of presynaptic actin dynamics for synapse function and mouse behavior. Exp. Cell Res. 2015, 335, 165–171.
- Bartol, T.M.; Bromer, C.; Kinney, J.; Chirillo, M.A.; Bourne, J.N.; Harris, K.M.; Sejnowski, T.J. Nanoconnectomic upper bound on the variability of synaptic plasticity. Elife 2015, 4, e10778.
- Bourne, J.N.; Chirillo, M.A.; Harris, K.M. Presynaptic ultrastructural plasticity along CA3→CA1 axons during long-term potentiation in mature hippocampus. J. Comp. Neurol. 2013, 521, 3898–3912.
- Meyer, D.; Bonhoeffer, T.; Scheuss, V. Balance and stability of synaptic structures during synaptic plasticity. Neuron 2014, 82, 430–443.
- Gundelfinger, E.D.; Fejtova, A. Molecular organization and plasticity of the cytomatrix at the active zone. Curr. Opin. Neurobiol. 2012, 22, 423–430.
- Matz, J.; Gilyan, A.; Kolar, A.; McCarvill, T.; Krueger, S.R. Rapid structural alterations of the active zone lead to sustained changes in neurotransmitter release. Proc. Natl. Acad. Sci. USA 2010, 107, 8836–8841.
- Monday, H.R.; Castillo, P.E. Closing the gap: Long-term presynaptic plasticity in brain function and disease. Curr. Opin. Neurobiol. 2017, 45, 106–112.
- Runge, K.; Cardoso, C.; de Chevigny, A. Dendritic Spine Plasticity: Function and Mechanisms. Front. Synaptic Neurosci. 2020, 12, 36.
- Nakahata, Y.; Yasuda, R. Plasticity of Spine Structure: Local Signaling, Translation and Cytoskeletal Reorganization. Front. Synaptic Neurosci. 2018, 10, 29.
- Holcman, D.; Triller, A. Modeling synaptic dynamics driven by receptor lateral diffusion. Biophys. J. 2006, 91, 2405–2415.
- Yang, X.; Specht, C.G. Subsynaptic Domains in Super-Resolution Microscopy: The Treachery of Images. Front. Mol. Neurosci. 2019, 12, 161.
- Hruska, M.; Henderson, N.; Le Marchand, S.J.; Jafri, H.; Dalva, M.B. Synaptic nanomodules underlie the organization and plasticity of spine synapses. Nat. Neurosci. 2018, 21, 671–682.
- Wegner, W.; Mott, A.C.; Grant, S.G.N.; Steffens, H.; Willig, K.I. In vivo STED microscopy visualizes PSD95 sub-structures and morphological changes over several hours in the mouse visual cortex. Sci. Rep. 2018, 8, 219.
- Moretto, E.; Longatti, A.; Murru, L.; Chamma, I.; Sessa, A.; Zapata, J.; Hosy, E.; Sainlos, M.; Saint-Pol, J.; Rubinstein, E.; et al. TSPAN5 Enriched Microdomains Provide a Platform for Dendritic Spine Maturation through Neuroligin-1 Clustering. Cell Rep. 2019, 29, 1130–1146.
- Choquet, D.; Triller, A. The role of receptor diffusion in the organization of the postsynaptic membrane. Nat. Rev. Neurosci. 2003, 4, 251–265.
- Triller, A.; Choquet, D. New concepts in synaptic biology derived from single-molecule imaging. Neuron 2008, 59, 359–374.
- Blanpied, T.A.; Scott, D.B.; Ehlers, M.D. Dynamics and regulation of clathrin coats at specialized endocytic zones of dendrites and spines. Neuron 2002, 36, 435–449.
- Rácz, B.; Blanpied, T.A.; Ehlers, M.D.; Weinberg, R.J. Lateral organization of endocytic machinery in dendritic spines. Nat. Neurosci. 2004, 7, 917–918.
- Lu, J.; Helton, T.D.; Blanpied, T.A.; Rácz, B.; Newpher, T.M.; Weinberg, R.J.; Ehlers, M.D. Postsynaptic positioning of endocytic zones and AMPA receptor cycling by physical coupling of dynamin-3 to Homer. Neuron 2007, 55, 874–889.
- Groc, L.; Choquet, D. Measurement and characteristics of neurotransmitter receptor surface trafficking (Review). Mol. Membr. Biol. 2008, 25, 344–352.
- Kusters, R.; van der Heijden, T.; Kaoui, B.; Harting, J.; Storm, C. Forced transport of deformable containers through narrow constrictions. Phys. Rev. E. Stat. Nonlin. Soft Matter Phys. 2014, 90, 33006.
- Wang, J.; Fourriere, L.; Gleeson, P.A. Local Secretory Trafficking Pathways in Neurons and the Role of Dendritic Golgi Outposts in Different Cell Models. Front. Mol. Neurosci. 2020, 13, 597391.
- Frotscher, M.; Studer, D.; Graber, W.; Chai, X.; Nestel, S.; Zhao, S. Fine structure of synapses on dendritic spines. Front. Neuroanat. 2014, 8, 94.