Visual plasticity is classically considered to occur essentially in the primary and secondary cortical areas. Subcortical visual areas such as the dorsal lateral geniculate nucleus (dLGN) or the superior colliculus (SC) have long been held as basic structures responsible for a stable and defined function. In this model, the dLGN was considered as a relay of visual information travelling from the retina to cortical areas and the SC as a sensory integrator orienting body movements towards visual targets. However, recent findings suggest that both dLGN and SC neurons express functional plasticity, adding unexplored layers of complexity to their previously attributed functions. The existence of neuronal plasticity at the level of visual subcortical areas redefines our approach of the visual system.
- superior colliculus
- synaptic plasticity
- lateral geniculate nucleus
- visual system
- functional plasticity
1. Introduction
1.1. Lateral Geniculate Nucleus and Superior Colliculus
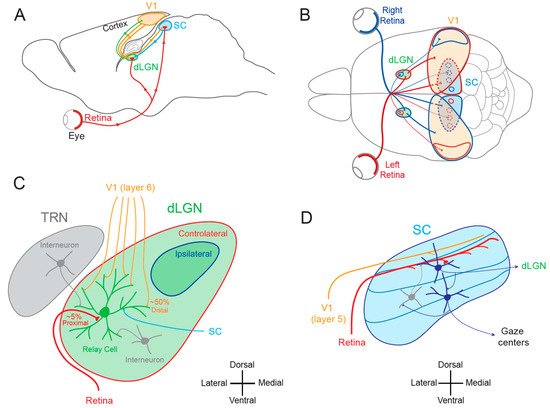
1.2. Cortical and Subcortical Plasticity
2. Functional Plasticity in Subcortical Visual Areas
2.1. Functional Plasticity in the dLGN
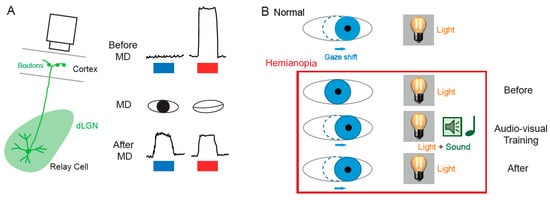
The notion that dLGN neurons do not express plasticity was disproved a decade after Wiesel and Hubel’s publication. Indeed, Ikeda and Wright showed in 1976 that the spatial resolution of dLGN neurons activated by the deviating eye in kittens reared with a squint is considerably reduced compared to that of neurons activated by the normal eye [18][28]. This result was the first to suggest that loss of normal binocular vision leads to plastic changes in the LGN. Later on, it was shown in amblyopic patients that functional deficits in visual response are already observed at the stage of the dLGN [19][29]. In addition, in contrast to what was previously assumed, about half of the rodent dLGN relay neurons in a given monocular territory in fact receive inputs from both eyes, indicating a potential binocularity for a large proportion of dLGN neurons [20][21][22][23][24][25][26][30,31,32,33,34,35,36]. Moreover, spatial receptive fields at eye opening in mouse dLGN are ~2 times larger than in adulthood due to an increase in surround suppression owing to an increased in feed-forward inhibition [27][37]. Furthermore, monocular deprivation (MD) in the mouse has been shown to produce a large shift in ocular dominance (OD) in dLGN neurons (Figure 2A) [28][29][30][38,39,40]. In one of these studies, GABAergic synaptic inhibition was found to be critical [29][39]. It is very unlikely that the plasticity observed in the dLGN only represents altered feedback from the cortex, because the shift in dLGN responses was resistant to cortical inactivation using the GABAA receptor agonist muscimol [28][38].
2.2. Functional Plasticity in the SC
2.2. Functional Plasticity in the SC
The SC is the mammalian equivalent of the optic tectum in inferior vertebrates [5]. While many studies reported functional and synaptic plasticity in the tadpole optic tectum [33][34][35][36][43,44,45,46], fewer investigations have been performed on the mammalian SC. As for the dLGN, the SC was thought to be largely devoid of functional plasticity, since receptive field features were found unchanged after MD [37][47]. However, recent findings suggest that SC express functional plasticity. The best demonstration of SC plasticity comes from studies on hemianopia, a permanent visual deficit caused by cortical trauma [31][32][38][39][41,42,48,49]. Patients with unilateral hemianopia are totally blind in the contralateral visual hemi-field but have preserved subcortical visual structures such as the SC. In basic post-traumatic conditions, hemianopia patients display a total lack of gaze orientation towards the blind hemi-field; a behavioral response depending on cortico-collicular connections. However, when the visual stimulus was temporally paired with an auditory stimulus occurring in the same region of the visual space (i.e., audio-visual training), normal gaze orientation towards the blind hemi-field (Figure 2B) was observed both in patients [31][41] and cats [32][39][42,49]. Interestingly, the re-emergence of visual behavior in cats is correlated with the reinstatement of visual responsiveness in deep layer neurons of the ipsilesional SC [32][42]. This audio-visual training procedure is thought to be related to the Hebbian learning and to reflect potentiation of visually activated synapses onto gaze-orientation related premotor neurons within the SC that fired under the conjoint activation of auditory synapses. In fact, audio-visual training has been shown to be able to reveal auditory or visual responses that were absent initially [40][50].
3. Structural Plasticity in Subcortical Visual Areas
3.1. Structural Plasticity in the dLGN
3.2. Structural Plasticity in the SC
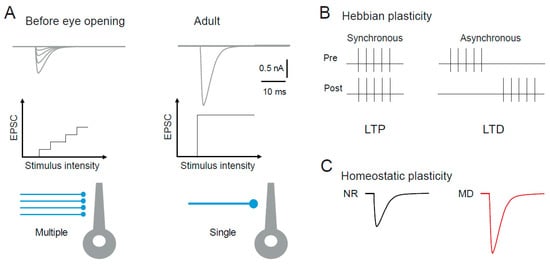
4. Synaptic Plasticity in Subcortical Visual Areas
4.1. Synaptic Plasticity in the dLGN
4.1.1. Hebbian Synaptic Plasticity in the dLGN
4.1.2. Homeostatic Synaptic Plasticity in the dLGN
4.2. Synaptic Plasticity in the SC
4.2.1. Hebbian Synaptic Plasticity in the SC
4.2.2. Homeostatic Synaptic Plasticity in the SC
5. Intrinsic Plasticity in Subcortical Visual Areas
Beyond synaptic plasticity, changes in intrinsic neuronal excitability represent the other side of functional plasticity that usually goes hand-in-hand with synaptic modifications [75][76][104,105] and possibly participate to developmental plasticity and learning [77][78][106,107]. Intrinsic plasticity is generally triggered by synaptic activity (induction phase) that induces plasticity of synaptic transmission in parallel. However, the expression of plasticity of intrinsic neuronal excitability depends on the regulation of voltage-gated ion channels (expression phase) such as hyperpolarization-activated cyclic nucleotide-gated (HCN) channels [79][80][81][82][108,109,110,111], Nav [83][112], Kv1 [84][113] and Kv7 [85][114] channels.