1. Introduction
Inflammation is a kind of active defense reaction of organisms to external stimulations, such as infectious microorganisms, or internal processes, such as tissue injury, cell death, and cancer
[1][2][3][1,2,3]. However, long-term low-grade inflammation leads to many human diseases, including aging, metabolic disorders, cancer, and neurodegenerative diseases
[4][5][6][7][4,5,6,7]. Thus, the discovery of anti-inflammatory medicines has been and is continuing to be one of the hotspots of pharmaceutical research.
Currently, anti-inflammatory therapy mainly includes non-steroidal anti-inflammatory drugs (NSAIDS) and glucocorticoids, both of which possess various side effects, such as cardiotoxicity, hepatotoxicity, and immunological dysfunction
[8][9][14,15]. Natural products have attracted increasingly more attention due to their safety and effectiveness
[10][16]. Emerging evidence indicates that natural products always function as multi-component and multi-target patterns
[11][17]. Naturally occurring anti-inflammatory compounds might be promising candidates for the treatment of enteritis, arthritis, and skin inflammation. Xanthones were firstly isolated in 1855 by a German scientist pursuing research on dysentery and then named by the Greek word for yellow, xanthos
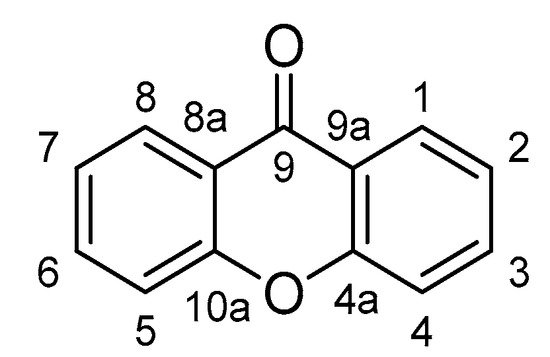
[12][18]. Xanthones possess a unique 9
H-Xanthen-9-one scaffold (
Figure 1), which mainly occurs in the plants of the families Gentianaceae and Hypericaceae, as well as some fungi and lichens
[13][19]. Several types of xanthones have been identified, including simple oxygenated xanthones, xanthone glycosides, prenylated xanthones, xanthonolignoids, and miscellaneous
[14][20]. The studies of xanthone are provoking not only due to the structural diversity but also a variety of pharmacological activities. Many xanthones have been reported with potent anti-inflammatory properties
[15][16][17][18][19][21,22,23,24,25].
Figure 1.
The core structure of xanthone.
2. Xanthones with Anti-Inflammatory Properties
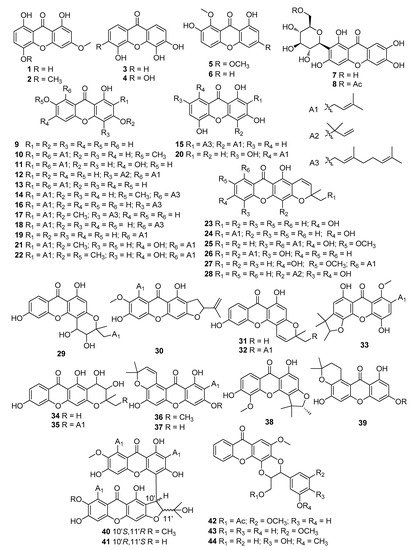
Using the keywords xanthone and inflammation, we collected data from Google Scholar, Web of Science, Scopus, and Pubmed. A total of 44 xanthones were found with anti-inflammatory properties, containing 6 simple oxygenated xanthones (1–6), 2 xanthone glycosides (7, 8), 33 prenylated xanthones (9–41), and 3 xanthonolignoids (42–44) (Figure 2). Many models, either in vitro or in vivo, have been recruited to evaluate the anti-inflammatory properties of xanthones. To organize the review, the xanthones were classified based on bioassays (Table 1).
Figure 2.
Structures of xanthones with anti-inflammatory activity. The symbol Ac represents an acetyl group.
Table 1. Xanthones with anti-inflammatory activity.
|
| Model/Method |
|
| No |
|
| Dose |
|
| Outcomes |
|
| Ref. |
|
|
| LPS-stimulated RAW264.7 macrophages |
|
| 1 |
|
| 10 μmol/L |
|
| Suppressed the phosphorylation of IKK-β, Akt, and p65 |
|
[20]
|
[38]
|
|
| 2 |
|
| 10 μmol/L |
|
| Inhibited the production of IL-6 and TNF-α |
|
[20]
|
[38]
|
|
| 4, 7 |
|
| 25, 50 μg/mL |
|
| Suppressed the generation of TNF-α and ICAM-1 |
|
[21]
|
[28]
|
|
| 6, 15−18, 21, 26−28, 37 |
|
| 3, 10, 30, 100 μmol/L |
|
| Downregulated mRNA expressions of iNOS and COX-2 |
|
[22]
|
[37]
|
|
| 9 |
|
| 50 μmol/L |
|
| Suppressed iNOS, COX-2, inhibited TNF-α, IL-1β, IL-6, IκB-α |
|
[23]
|
[34]
|
|
| 12 |
|
| 1, 2, 5, 10 μmol/L |
|
| Induced HO-1 expression and increased HO-1 activity, inhibited TNF-α, IL-1β |
|
[24]
|
[35]
|
|
| 19 |
|
| 5, 10, 20 μmol/L |
|
| Inhibited NO production and IL-6 secretion |
|
[25]
|
[29]
|
|
| 22 |
|
| 11.72 ± 1.16 μmol/L |
|
| Inhibited NO production |
|
[26]
|
[30]
|
|
| 30 |
|
| 20, 40, 60 μmol/L |
|
| Inhibited the production of NO, iNOS, TNF-α, IL-6, and IL-1β |
|
[27]
|
[31]
|
|
| 33 |
|
| 6.25 μmol/L |
|
| Suppressed NO production |
|
[28]
|
[32]
|
|
| 38 |
|
| 50 μg/mL |
|
| Inhibited COX-1, COX-2 and 5-LOX-mediated LTB4 formation |
|
[29]
|
[36]
|
|
| 40 |
|
| 11.3 ± 1.7 μmol/L |
|
| Inhibited NO production |
|
[30]
|
[33]
|
|
| 41 |
|
| 18.0 ± 1.8 μmol/L |
|
| Inhibited NO production |
|
[30]
|
[33]
|
|
| LPS/IFN?-stimulated RAW264.7 macrophages |
|
| 20 |
|
| 3.125–25 ?mol/L |
|
| Suppressed IL-6, IL-12, and TNF-? |
|
[31]
|
[69]
|
|
| 39 |
|
| 10 μmol/L |
|
| Decreased NO production |
|
[32]
|
[40]
|
|
| Human neutrophils |
|
| 3, 7, 42, 43 |
|
| 1000 μg/mL |
|
| Inhibited WST-1 by NADPH oxidase |
|
[33]
|
[45]
|
|
| 23, 24, 29, 31, 32, 34, 35 |
|
| 10 μg/mL |
|
| Inhibited superoxide anion generation and elastase release |
|
[34]
|
[44]
|
|
| CD3− synovial cells |
|
| 7 |
|
| 100 μg/mL |
|
| Downregulation of TNF-α, IL-1β, and IFN-γ |
|
[35]
|
[46]
|
|
| Lung of septic mice |
|
| 10, 30, 100 mg/kg |
|
| Upregulated the expression and activity of HO-1 |
|
[36]
|
[48]
|
|
| Carrageenan-induced mechanical hyperalgesia Wistar rats |
|
| 100 μg/paw |
|
| Inhibited TNF-α level through CINC-1/epinephrine/PKA pathway |
|
[37]
|
[51]
|
|
| MC 3T3-E1 cell line |
|
| 10, 20, 30, 40 μmol/L |
|
| Alleviated oxidative stress by activating the BMP2/Smad-1 signaling pathway |
|
[38]
|
[54]
|
|
| HFLS-RA cells |
|
| 10 |
|
| 10 μg/mL |
|
| Inhibited nuclear translocation of p65 |
|
[39]
|
[56]
|
|
| AA rats |
|
| 10 |
|
| 2.5−10 μg/mL |
|
| Inhibited fibrous hyperplasia, synovial angiogenesis, cartilage |
|
[39]
|
[56]
|
|
| Peripheral LPS-induced neuroinflammation in C57BL/6J mice |
|
| 10 |
|
| 40 mg/kg |
|
| Reduced brain levels of IL-6 and COX-2 |
|
[40]
|
[58]
|
|
| Established CIA in DBA/1J mice |
|
| 10 |
|
| 10, 40 mg/kg |
|
| Reduced the levels of anti-collagen IgG2a and autoantibodies in serum and the production of LIX/CXCL5, IP-10/CXCL10, MIG/CXCL9, RANTES/CCL5, IL-6 and IL-33 in joints |
|
[41]
|
[60]
|
|
| Ovalbumin-induced allergic asthma mice |
|
| 9, 10 |
|
| 10, 30 mg/kg |
|
| Increased Th2 cytokine |
|
[42]
|
[62]
|
|
| 3T3-L1 cells |
|
| 10, 19 |
|
| 10 μmol/L |
|
| Inhibited PPARγ and NFR2 through NF-κB |
|
[43]
|
[64]
|
|
| Acetic acid-induced mice |
|
| 5 |
|
| 10, 20 mg/kg |
|
| Reduced paw edema |
|
[44]
|
[65]
|
|
| EPP-induced ear edema |
|
| 10, 13, 14, 25, 36 |
|
| 1 mg/kg |
|
| Inhibited edema |
|
[45]
|
[67]
|
|
| LPS-induced adipose tissue inflammation mice |
|
| 10 |
|
| 10 mg/kg |
|
| Reduced macrophage content and shifted pro-inflammatory macrophage polarization |
|
[12]
|
[18]
|
|
| 19 |
|
| 20 mg/kg |
|
| Reduced macrophage content through inhibiting MAPKs and NF-κB activation |
|
[25]
|
[29]
|
Macrophages are a major component of the mononuclear phagocyte system
[46][26]. Monocytes migrate into various tissues and transform into macrophages. Macrophages play a critical role in the initiation, maintenance, and resolution of inflammation. Lipopolysaccharide (LPS), a component of the Gram-negative bacterial cell wall, has been widely used to induce an inflammatory response in macrophages
[47][27]. The LPS-stimulated RAW264.7 macrophage model is an effective tool for anti-inflammatory drug screening and anti-inflammatory mechanism investigation. Using a LPS-induced RAW264.7 macrophage model, 3,4,5,6-tetrahydroxyxanthone (
4) and mangiferin (
7) were found to suppress the generation of TNF-α and intercellular adhesion molecule-1 (ICAM-1)
[21][28]. Six xanthones, including 1,3,6,7-tetrahydroxy-8-prenylxanthone (
19)
[25][29], β-mangostin (
22)
[26][30], nagostenone F (
30)
[27][31], inophinnin (
33)
[28][32], garcinoxanghone B (
40)
[30][33], and garcinoxanthone C (
41)
[30][33], were reported to reduce NO production in LPS-stimulated RAW264.7 macrophage. 6′-
O-acetyl mangiferin (OAM) (
8), an acetylated xanthone C-glucoside, was reported to suppress iNOS and COX-2 expression, thereby inhibiting the levels of TNF-α, IL-1β, and IL-6 in LPS-stimulated RAW264.7 cells. Furthermore, OAM inhibited the LPS-induced phosphorylation of c-Jun N-terminal kinases (JNK), extracellular signal-regulated kinase (ERK), and p38, which led to the blockade of nuclear factor-κB (NF-κB) and inhibitor κB (IκB)-α activation
[23][34]. Cudratricusxanthone A (
12), isolated from the roots of
Cudrania tricuspidata Bureau (Moraceae), was found to induce heme oxygenase-1 (HO-1) expression at a non-cytotoxic concentration (1–10 μmol/L) in LPS-treated RAW264.7 macrophages, which in turn suppressed PGE2, NO, TNF-?, and IL-1β production
[24][35]. Dual arachidonate 5-lipoxygenase (5-LOX) and COX inhibitors are potential new drugs to treat inflammation. 6-Dihydroxy-5-methoxy-4′,5′-dihydro-4′,4′,5′-trimethylfurano-(2′,3′:3,4)-xanthone (
38) inhibited COX-1 and COX-2 production, and 5-LOX mediated leukotriene B4 (LTB4) formation in LPS-induced RAW264.7 macrophages
[29][36]. Among the xanthones isolated from the roots of
Cratoxylum formosum ssp.
pruniflorum, several compounds (
6, 1
5–
18,
21,
26–
28, and
37) showed anti-inflammatory activity in LPS-induced RAW 264.7 macrophages
[22][37]. Dulxisxanthone F (
21) was found to downregulate the mRNA expression of iNOS and COX-2 in dose-dependent manners, and 5,9-dihydroxy-8-methoxy-2,2-dimethyl-7-(3-methylbut-2-enyl)-2
H,6
H-pyrano-[3,2
b]-xanthone (
27) only inhibited the mRNA expression of iNOS but not COX-2. Two xanthones, ellidifolin (
1) and swerchirin (
2), were isolated from
Swertia chiraytia [20][38], which were found to inhibit the production of pro-inflammatory cytokines IL-6 and TNF-? in LPS-stimulated RAW264.7 macrophages; furthermore, ellidifolin (
1) inhibited the production of PGE2 by suppressing the phosphorylation of JNK, ERK, and p38 MAPKs (mitogen-activated protein kinases).
Interferon γ (IFN?) is a dimerized soluble cytokine, and aberrant IFN? expression is related to a number of inflammatory and autoimmune diseases
[48][39]. LPS plus IFN? stimulation caused the increase of TNF receptor associated factor family member-associated NF-κB activator binding kinase 1 (TBK1) expression, p50/p65 nuclear translocation, and activation of NF-κB in RAW264.7 macrophages, 1,3,5,7-tetrahydroxy-8-prenylxanthone (
20), reversed the above changes to suppress the production of IL-6, IL-12, and TNF-?
[32][40]. In IFN? plus LPS-induced RAW264.7 macrophages, hyperxanthone E (
39) was reported to decrease NO production
[32][40].
The major role of neutrophils in the host defense is to eliminate invading microorganisms
[49][41]. In neutrophils,
N-formylmethionyl-leucyl-phenylalanine (fMLP) is a powerful activator of polymorphonuclear and mononuclear phagocytes, and the effects of fMLP on neutrophil activity can be inhibited by pertussis toxin
[50][42]. The neutrophil-mediated inflammatory response is regarded as a multi-step process involving the initial adhesion of circulating neutrophils to activated vascular endothelium
[51][43]. In fMLP/CB-stimulated human neutrophils, several gambogic acid analogs (
23,
24,
29,
31,
32,
34, and
35) inhibited superoxide anion generation and elastase release
[34][44]. Several xanthons (
3,
9,
42, and
43) were isolated from the twigs of
Hypericum oblongifolium wall, which showed anti-inflammatory activity in isolated human neutrophils
[33][45].
CD3
− synovial cells are suggested to play an important role in RA development and therefore are a perfect model in the search for new anti-arthritic drugs. Mangiferin (
7) downregulated TNF-α, IL-1β, and IFN-γ expression in TNF-α-stimulated CD3
− synovial cells from rheumatoid arthritis (RA) patients, which indicated that mangiferin could be a potent candidate for the treatment of RA
[35][46].
Sepsis is a major cause of death worldwide
[52][47]. Infection-induced inflammation is strongly regulated by many endogenous negative feedback mechanisms that modulate the intensity of inflammation, promote its eventual resolution, and return it back to homeostasis. Mangiferin (
7) dose-dependently upregulated the expression and activity of HO-1 in the lung from septic mice
[36][48].
Carrageenan is a pro-inflammatory agent used as a tool to induce inflammatory hyperalgesia in rats and mice
[53][49]. The carrageenan-induced peripheral inflammatory pain model is widely used because it resembles inflammatory pain susceptible to both steroidal and nonsteroidal anti-inflammatory drugs
[54][50]. Local administration of mangiferin (
7) prevented inflammatory mechanical hyperalgesia induced by carrageenan in rats, which depended on the inhibition of TNF-α production/release and the CINC1 (cytokine-induced neutrophil chemoattractant 1)/epinephrine/PKA (protein kinase A) pathway
[37][51].
MC3T3 is an osteoblast precursor cell line derived from
Mus musculus (mouse), which is one of the most convenient and physiologically relevant systems for the study of transcriptional control in calvarial osteoblasts
[55][52]. Dexamethasone is a known synthetic glucocorticoid, which induces sodium-dependent vitamin C transporter in MC3T3-E1 cells
[56][53]. Bone morphogenetic protein 2 (BMP2) plays a role in postnatal bone formation, mediated by activating ligand-bound Small Mothers Against Decapentaplegic (SMAD) family members
[20][38]. Mangiferin (
7) attenuated dexamethasone-induced injury and inflammation in MC3T3-E1 cells by activating the BMP2/Smad-1 signaling pathway
[38][54].
HFLS-RA is a human fibroblast-like synoviocyte with high proliferating ability and susceptibility. HFLS-RA cell is an excellent cellular model for studying synoviocyte physiology in relation to the development and treatment of RA
[57][55]. α-Mangostin (
10) (10 μg/mL) was found to suppress the expression and activation of key proteins in the NF-κB pathway and inhibit the nuclear translocation of p65 in HFLS-RA cells
[39][56].
Adjuvant-induced arthritis (AA) is evaluated by paw edema, arthritis score, and hematological parameters. α-Mangostin (
10) protected joints from rats suffering from AA, indicated by attenuated paw swelling, reduced inflammatory cell infiltration, decreased secretion of IL-1β and TNF-α in serum, and inhibition of NF-κB activation in synovia
[39][56].
The presence of neuroinflammation is a common feature of dementia
[58][57]. Reactive microgliosis, oxidative damage, and mitochondrial dysfunction are associated with the pathogenesis of all types of neurodegenerative dementia, such as Parkinson’s disease dementia (PDD), frontotemporal dementia (FTD), Alzheimer’s disease (AD), and Lewy body dementia (LBD). Peripheral LPS-induced neuroinflammation in C57bl/6J mice has been used to evaluate neuroinflammation and neurodegeneration as an adjuvant therapeutic strategy. α-Mangostin (
10) reduced the levels of proinflammatory cytokine IL-6, COX-2, and 18 kDa translocator protein (TSPO) in the brain from LPS-induced neuroinflammation in C57BL/6J mice, which was considered as an adjuvant treatment in preclinical models of AD, PD, and multiple sclerosis
[40][58].
RA is a long-term autoimmune disease in which the body’s immune system mistakenly attacks the joints; RA causes pain, stiffness, and swelling in the joints
[59]. α-Mangostin (
10) decreased the clinical score at both doses (10 and 40 mg/kg) and decreased the histopathological score at the high dose in collagen-induced arthritis (CIA) in DBA/1J mice
[41][60].
Asthma is a chronic inflammatory disease of the airways characterized by reversible airway obstruction, airway hyperreactivity (AHR), and remodeling of the airways
[60][61]. Allergic asthma is associated with excessive T helper type 2 (Th 2) cell activation and AHR
[57][55].