Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Alvaro Compañ-Bertomeu and Version 2 by Jessie Wu.
Mesenchymal stem/stromal cells (MSCs) represent a promising therapy for musculoskeletal diseases. There is compelling evidence indicating that MSC effects are mainly mediated by paracrine mechanisms and in particular by the secretion of extracellular vesicles (EVs). Many studies have thus suggested that EVs may be an alternative to cell therapy with MSCs in tissue repair.
- mesenchymal stem cells
- extracellular vesicles
- immunoregulation
- bone diseases
- osteoarthritis
1. Introduction
Musculoskeletal disorders and injuries are the leading cause of disability worldwide with a high prevalence across the life-course. People with multi-morbidity are particularly vulnerable especially in the context of an aging population. These disorders are painful and lead to mobility limitation and early retirement with a high economic and social impact. They include a wide range of conditions such as osteopenia, osteoporosis, fractures, osteoarthritis (OA), rheumatoid arthritis (RA), sarcopenia and so forth [1].
Transplantation of mesenchymal stem/stromal cells (MSCs) has been recognized in recent years as a promising therapy for musculoskeletal diseases. Preclinical models have provided evidence that MSCs have potential applications in these conditions due to their regenerative and immunomodulatory properties. MSC exhibit a variety of trophic activities relevant to musculoskeletal therapy (reviewed in Reference [2]). The efficacy of MSC treatment has been demonstrated in animal models and clinical studies of RA, OA and cartilage repair [3][4][5][6][7][3,4,5,6,7], as well as in bone [8][9][8,9], tendon [10] and skeletal muscle [11] regeneration. Although more research is needed including controlled clinical studies with long-term follow-up, these results open the possibility of improving current therapies.
A wide range of evidence has demonstrated that paracrine mechanisms are main components of MSC regenerative effects. MSCs respond to stimuli present in the local microenvironment by secreting a variety of bioactive molecules. Accordingly, it is possible to modulate the composition of the MSC secretome by cellular pre-conditioning during culture, thus maximizing their potential for therapeutic applications (reviewed in Reference [12]). The conditioned medium (CM) from MSCs contains multiple factors that may cooperate to induce a repair response. Besides extracellular vesicles (EVs), MSCs can secrete a wide range of molecules such as purines, bone morphogenetic proteins (BMPs), CD274, C-C motif chemokine ligand (CCL)-2, connexin 43, indoleamine 2,3-dioxygenase, prostaglandin (PG)E2, interleukin(IL)-6, IL-10, NO, heme oxygenase-1, tumor necrosis factor-inducible gene-6 (TSG-6), leukemia inhibitory factor (LIF), CD95/CD95 ligand, galectins, human leukocyte antigen-G5 (HLA-G5) and growth factors such as transforming growth factor-β1 (TGF-β1), hepatic growth factor (HGF), vascular endothelial growth factor (VEGF), platelet-derived growth factor, fibroblast growth factor (FGF) and so forth. [2][13][14][15][16][17][18][19][2,13,14,15,16,17,18,19]. The use of CM may avoid some problems associated with the therapeutic application of MSCs such as immune rejection of allogeneic cells or undesirable cell differentiation. However, treatment with CM may be an alternative to cellular therapy in regenerative medicine [20].
EVs are main components of MSC secretome which can be incorporated into cells via endocytosis or phagocytosis [21][22][23][21,22,23] leading to the transfer of their content such as proteins, lipids, DNA, RNA, mitochondria and so forth. As a result, EVs regulate gene transcription and the functions of recipient cells [24][25][26][27][24,25,26,27]. In particular, the transfer of miRNA plays an important role in the biological activity of MSC EVs [28]. Most of the bioactive effects of EVs require the interaction between EV-associated molecules CD9 and CD81 and the binding partners immunoglobulin superfamily, member 8 (IGSF8) and PGF2 receptor negative regulator (PTGFRN) on cells [24]. It is widely accepted that EVs represent an important mechanism for cell communication.
EVs are generally classified into exosomes (EXOs, 30–100 nm in diameter, formed by the inward budding of endosomal membrane to produce a multivesicular body that upon fusion with cell membrane releases these microparticles) and microvesicles (MVs, 50 nm to 1000 nm in diameter, generated by the outward budding and fission of the plasma membrane) (reviewed in Reference [29]). Nevertheless, MSCs may secrete other EVs with overlapping size which do not exhibit the same characteristics as EXOs [30]. As currently there are no appropriate methods to isolate pure EV subtypes, better isolation and characterization methods are under study to establish the necessary protocols and classifications needed to know their specific properties allowing the development of EVs for therapeutic applications. To this respect, the International Society for Extracellular Vesicles (ISEV) is committed to improving the standardization guidelines for EV studies [31].
2. Immune Regulation and Rheumatoid Arthritis
A wide range of evidence indicates that EVs play an essential role in immunomodulation. EVs may either activate or suppress immune responses depending on the type of parent cell [32][33][36,37]. However, many in vitro and in vivo studies have demonstrated the predominant anti-inflammatory and immunosuppressive properties of MSC EVs in both innate and adaptive immune cells [34][35][38,39]. These therapeutic effects of EVs may be similar to those of parent cells [36][40]. EVs contain a number of tolerogenic molecules present in MSCs (programmed death-ligand 1, galectin-1 and TGF-β1) [37][41]. Nevertheless, some studies have reported a higher immunomodulatory ability of MSCs compared with their EVs which may be mediated by cell contact besides a number of factors present in the secretome [38][42]. Another study has also shown that B cell modulation by MSCs is partially mediated by soluble factors other than EVs [39][43]. It has been reported that EVs from human bone marrow MSCs (BM-MSCs) reduce the expression of pro-inflammatory cytokines such as IL-1β, IL-6 and tumor necrosis-α (TNFα) in monocytic cells whereas the expression of anti-inflammatory IL-10 and TGF-β1 is enhanced [40][44]. The anti-inflammatory properties of EVs can be potentiated by stimulation of MSCs with cytokines such as TNFα+interferon γ [41][45].
MSC EVs exert immunoregulatory effects by several mechanisms (Figure 1). EVs regulate dendritic cell functions with impairment of antigen uptake by immature cells and reductions in cell maturation and activation [42][46]. Although there are differences depending on the parent cells, MSC EVs may inhibit the proliferation of CD4+ and CD8+ T cells and the promotion of apoptosis in CD4+ T cells [43][44][47,48]. EVs may also promote the conversion of T helper type 1 (Th1) into T helper type 2 (Th2) cells whereas differentiation into Th17 cells was decreased [40][44]. In addition, EVs polarize activated CD4+ T cells to CD4+CD25+FOXP3+ regulatory T cells (Tregs) which requires activation of T cells by antigen presenting cells [43][45][46][47,49,50]. As MSC EVs polarize immune cells toward an immunosuppressive phenotype only in the presence of an activated immune system [46][50], homeostatic immune activity would not be affected by EVs treatment thus avoiding the increased risk of infection or cancer observed with immunosuppressive drugs [47][51]. The proliferation and differentiation of B cells and the proliferation of natural killer (NK) cells are reduced by MSC EVs [48][49][52,53] while the levels of cytotoxic T lymphocyte-associated protein 4 (CTLA-4), a protein receptor which downregulates immune responses, are enhanced [40][44]. In particular, EVs have the potential to switch macrophages into an anti-inflammatory phenotype (M2) [50][54]. Cell pre-conditioning can improve the macrophage differentiation ability of EVs as shown by using hypoxia which increases the content of miRNAs such as miR-223 and miR-146b in EVs [51][55]. Similarly, treatment of human umbilical cord MSCs (UC-MSCs) with lipopolysaccharide (LPS) has been shown to improve the anti-inflammatory ability of their EXOs through the induction of let-7b which is transferred to recipient cells and targets the toll-like receptor 4 (TLR4)/nuclear factor κ-light-chain-enhancer of activated B cells (NF-κB)/signal transducer and activator of transcription 3 (STAT3)/Akt pathway. As a result, macrophages can be polarized into an anti-inflammatory M2 phenotype [52][56].
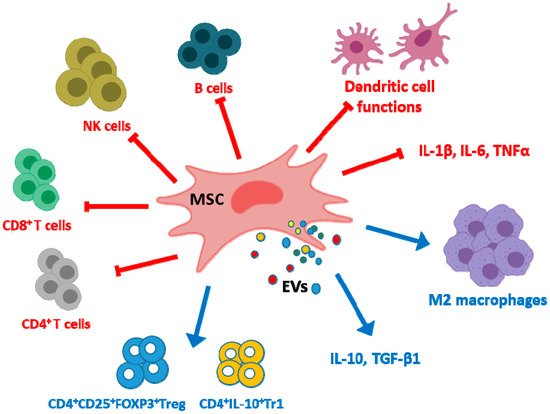
Figure 1. Examples of immunomodulatory effects exerted by mesenchymal stem/stromal cell extracellular vesicles (MSC EVs). EVs inhibit the production of pro-inflammatory cytokines but increase the levels of anti-inflammatory cytokines in monocytic cells. They exert negative effects on dendritic cell maturation and activation with impairment of antigen uptake and also on the proliferation of CD4+ T cells, CD8+ T cells, NK cells and B cells. These EVs promote the conversion of Th1 into Th2 and reduce Th17 differentiation. In contrast, MSC EVs enhance the differentiation of regulatory cells such as M2 macrophages, CD4+CD25+FOXP3+Treg and CD4+IL-10+Tr1.
RA is a chronic autoimmune inflammatory disorder characterized by hyperplasia of the synovial membrane and infiltration of immune and inflammatory cells. Synovial cell transformation, excessive production of inflammatory and catabolic mediators and osteoclast generation progressively result in joint destruction and disability [53][54][57,58]. Although there is limited information, EVs may represent a novel therapeutic strategy for arthritis. Experimental evidence indicates an immunomodulatory role for MSC EVs which may counteract antigen-driven T cell responses thus suggesting a potential interest of this approach for the treatment of T cell-mediated diseases such as RA. In a model of antigen (bovine serum albumin)-induced synovitis in pigs, intra-articular injection of EVs from pig BM-MSCs into the carpal joint (500 μg protein/injection in 500 μL) significantly reduced lymphocyte counts and gene expression of TNFα in synovial fluid. In addition, gait analysis at day 7 revealed a trend for the improvement of the impulse which may be due to pain reduction linked to the anti-inflammatory effect of EVs [55][59].
MVs and EXOs from BM-MSCs isolated from C57BL/6 mice did not reduce the proliferation of CD8+ or CD4+ T lymphocytes but increased CD4+CD25+FOXP3+ Tregs and the CD4+IL10+ Tr1 regulatory cell population. In the in vivo models of delayed-type hypersensitivity and collagen-induced arthritis (CIA) in mice both EV fractions exerted anti-inflammatory effects and protected joints from degradation, with a higher efficacy of EXOs, which may be related to the inhibition of plasmablast differentiation and the induction of IL-10-expressing regulatory B cells [56][60].
Annexin A1 is an endogenous inhibitory mediator of arthritis [57][61] enriched in synovial fluid neutrophil EVs from RA patients [58][62]. Animal models of RA have shown the role of annexin A1 in the anti-inflammatory and chondroprotective effects of neutrophil-derived EVs which depend on the interaction with its receptor formyl peptide receptor 2 (FPR2), inducing anabolic responses with TGF-β1 production and deposition of extracellular matrix, as well as chondrocyte protection from apoptosis [58][62]. Interestingly, annexin A1 is a main component of adipose tissue-derived mesenchymal stem cells (AD-MSCs) MVs and may contribute to the anti-inflammatory properties of these EVs [59][63].
3. Bone Repair and Bone Diseases
The secretome of MSCs mediates the mitogenic, pro-migratory and pro-osteogenic effects of these cells promoting bone healing in animal models through the recruitment of MSCs, endothelial cells and progenitor cells [24][60][24,64]. It has been reported that EXOs from AD-MSCs enhance the proliferation, migration and differentiation of human osteoblastic cells. These functions are potentiated by TNFα pre-conditioning of AD-MSCs which increases the expression of Wnt-3a in EXOs [61][65]. BM-MSC EVs promote osteoblast differentiation and expression of osteogenic genes, which is partly mediated by miR-196a as it has been demonstrated in functional experiments [62][66]. The differentiation stage of progenitor MSCs may influence the miRNA composition of EVs and their ability to induce osteogenic differentiation and mineralization in recipient MSCs [63][67].
Tissue regeneration requires the recruitment of endogenous stem/progenitor cells to the target site to form tissue specific cells as well as new vessels to provide the necessary oxygen, nutrients and growth factors to repair the affected area [64][65][68,69]. The repair effects of EVs on bone have been demonstrated both in vitro and in vivo, in different models of fracture, bone defect or osteoporosis. The relevance of MSC EXOs in bone healing has been shown in a femur fracture model of CD9-/- mice which have impaired EV formation. The retardation of callus formation and fracture healing can be rescued by the injection of BM-MSC EXOs but not by EXOs-free CM [66][70]. EV administration accelerates bone repair in models of calvarial bone defect in mice and rats. Thus, EVs enhanced osteoblastic proliferation at bone defect edge and stromal progenitor cell migration into the defect mid-substance leading to and increased re-ossification of the defect with increases in bone volume, bone formation area and healing score [24][62][24,66]. In addition, EVs from AD-MSCs combined with poly(lactic-co-glycolic acid) (PLGA) scaffolds have demonstrated an enhancing effect on osteogenic, proliferation and migration capabilities of human BM-MSCs in vitro. This approach also enhanced bone regeneration through these osteoinductive effects in critical-sized calvarial defect in mice [67][71]. Human induced pluripotent stem cells (iPSCs) obtained via genetic reprogramming of adult somatic cells may have some advantages compared with adult MSCs for therapeutic applications. Autologous iPSCs-derived MSCs (iMSCs) have great potential in regenerative medicine and they are superior to MSCs in cell proliferation ability, immunomodulation and secretion of bioactive factors including EVs [68][69][72,73]. Therefore, EXOs derived from human iMSCs combined with tricalcium phosphate (β-TCP) improved the osteoinductivity of these scaffolds through activation of phosphatidylinositol-3-kinase (PI3K)/Akt signaling in human BM-MSCs [70][74].
In bone tissue repair, MSC EVs promote osteogenesis and angiogenesis (Figure 2) which may depend on interactions with different cell types such as BM-MSCs and endothelial cells. EVs from MSCs can be useful tools in bone regenerative medicine as they induce lineage specific differentiation of BM-MSCs and can bind to extracellular matrix proteins such as type I collagen and fibronectin [71][75]. Nevertheless, the source of EVs influences these regenerative properties. Thus, EXOs from diabetic rat BM-MSCs showed a reduced effect on the osteogenic differentiation of BM-MSCs and the angiogenic activity of human umbilical cord endothelial cells, compared with EXOs from healthy rats. In addition, bone regeneration and neovascularization effects in rat calvarial defects were lower for EXOs from diabetic rats [72][76].
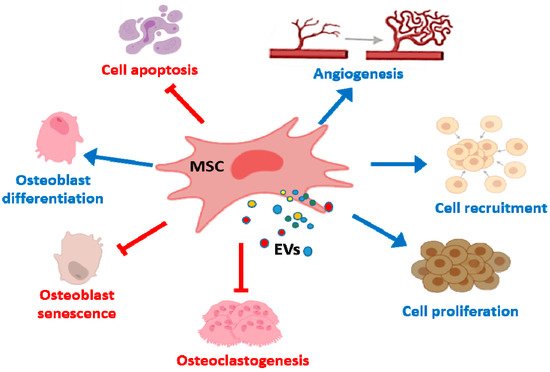
Figure 2. Effects of MSC EVs on bone repair and bone diseases. MSC EVs enhance proliferation, migration and osteogenic differentiation of BM-MSCs whereas apoptosis is inhibited. In addition, proliferation and recruitment of endothelial cells/endothelial progenitor are promoted leading to angiogenesis. MSC EVs also reduce osteoclast formation and osteocyte-like cells apoptosis.
EVs may be provide novel opportunities in endochondral repair of large bone defects which need bone grafts and costly interventions (reviewed in Reference [73][77]). Thus, EVs may be a better approach compared with cell-based therapies as implanted cells compete with endogenous progenitor cells for oxygen and nutrients in this ischemic microenvironment. In particular, MSCs cannot adapt their glucose consumption and do not possess the necessary glycolytic reserves to maintain their metabolism [74][78]. In fact, reduction of MSCs metabolic needs by induction of quiescence can enhance their survival under ischemic conditions in the lesion site [75][79] while hypoxia can reduce their osteogenic potential [76][80]. In addition, most culture-expanded MSCs cannot adapt to the microenvironment and they die or are phagocytosed by macrophages shortly after implantation in critical-sized bone defects. Distraction osteogenesis induces bone tissue regeneration in large bone defects through the recruitment of endogenous MSCs and endothelial cells/endothelial progenitor cells to the distraction gap. Administration of BM-MSC CM is an effective procedure to accelerate this process as it promotes the migration of Sca-1+/PDGFR-α+ BM-MSCs which differentiate into osteoblasts and CD31+ cells leading to angiogenesis in a mouse model. Soluble factors CCL-2, CCL-5 and CCL-7 can recruit bone marrow mononuclear cells while IL-3 and IL-6 recruit endothelial cells and progenitor cells to enhance osteogenic activity [60][64]. Administration of EVs from different origins may be another strategy to shorten these lengthy procedures. Recently, it has been demonstrated a role for EXOs from progenitor endothelial cells in the stimulation of angiogenesis thus accelerating bone regeneration during distraction osteogenesis [77][81].
Besides EVs from BM-MSCs and AD-MSCs, EVs from different MSCs are of interest in bone repair. Therefore, EXOs derived from UC-MSCs enhance fracture healing by promoting angiogenesis in a rat model of stabilized fracture. These EXOs are taken up by endothelial cells leading to hypoxia-inducible factor-1α (HIF-1α) activation and VEGF production with the result of cell proliferation, migration and tube formation [78][82].
Experimental evidence also indicates the potential application of MSC EVs in osteoporosis. To this respect, implantation of EXOs secreted by iMSCs incorporated in β-TCP scaffolds into critical size calvarial bone defects enhanced osteogenesis and angiogenesis in ovariectomized rats, a model of postmenopausal osteoporosis. In vitro experiments demonstrated the ability of these EVs to stimulate the proliferation and osteogenic differentiation of BM-MSCs from ovariectomized rats [79][83]. Autologous EVs from human urine-derived stem cells are a potential therapeutic agent for osteoporosis. Systemic injection of these EVs have been shown to promote osteogenesis and inhibit osteoclastogenesis in osteoporotic mice by transferring collagen triple-helix repeat containing 1 (CTHRC1) and osteoprotegerin proteins [80][84].
Senile osteoporosis may be another potential application of EVs. Therefore, bone loss was attenuated by i.v. administration of EVs from human umbilical cord blood plasma to 16 months old male C57BL/6 mice. This treatment increased trabecular and cortical bone mass and promoted osteoblast differentiation while reducing osteoclast formation. miR-3960 was enriched in these EVs and mediates their osteogenic effects [81][85]. Increased osteocyte apoptosis and osteoclast activation leads to imbalanced bone remodeling during ageing. AD-MSC EVs reduced hypoxia/serum deprivation-induced apoptosis of osteocyte-like cells through upregulation of Bcl-2/Bax and downregulation of reactive oxygen species (ROS), cytochrome c and activation of caspase-9 and caspase-3. In addition, these EVs reduced osteoclastogenesis suggesting the potential of AD-MSC EVs to improve age-related bone loss [82][86].
Apoptosis of bone marrow cells plays an important role in the destruction of bone tissue during long-term or high dose glucocorticoid treatment leading to osteonecrosis of the femoral head. Strategies able to prevent apoptosis and promote proliferation of bone marrow cells may be useful to start tissue repair and inhibit disease progression. One of these strategies is the application of MSCs or EVs. Therefore, EXOs derived from human synovial MSCs are internalized by bone marrow cells to promote cell proliferation and inhibition of apoptosis. In a rat model of osteonecrosis of the femoral head induced by methylprednisolone, i.v. administration of 1 × 1011 particles of EXOs (in 200 μL) exerted a preventive effect and counteracted the inhibition of the osteogenic response induced by glucocorticoid [83][87]. It is known that EXOs secreted by human iMSCs promote angiogenesis due to the activation of PI3K/Akt signaling in endothelial cells. As a consequence, i.v. administration in the steroid-induced rat osteonecrosis model prevented bone loss and osteonecrosis of the femoral head [84][88]. As HIF-1α plays a role in osteogenesis and angiogenesis [85][86][89,90], strategies directed at the modulation of this transcription factor may improve the bone repair properties of EVs. Therefore, BM-MSCs transfected with a mutant HIF-1α which is not degraded in normoxic conditions were used to obtain mutant EVs which exhibited enhanced osteogenesis and angiogenesis properties leading to the repair of steroid-induced avascular necrosis of the femoral head in rabbits [87][91].
Potential therapeutic applications of MSCs from a variety of sources have been extended to periodontal defects [88][92]. Periodontitis leads to pathologic loss of periodontal ligament, cementum and alveolar bone [89][93]. Receptor activator of NF-κB ligand (RANKL) plays a main role in osteoclastogenesis during inflammatory bone resorption and activated T and B cells can be the source of this cytokine in periodontal disease [90][94]. MSCs from different origins have shown a periodontal regenerative potential in animal models without adverse effects. In particular, local implantation of periodontal ligament-derived MSCs have demonstrated their ability in the regeneration of periodontal ligament and cementum [88][92]. Similarly, collagen sponges containing CM from BM-MSCs have been shown to repair periodontal defects in rats 4 weeks after implantation with regeneration of alveolar bone, cementum and periodontal ligament. This CM contains a number of growth factors such as insulin-like growth factor-1, VEGF, TGF-β1 and HGF which could mediate mobilization of endogenous MSCs, angiogenesis and differentiation [91][95]. Interestingly, CM from BM-MSC stimulates alveolar bone augmentation prior to dental implant placement in humans without inducing an inflammatory response [92][96]. Implants of collagen sponges including EXOs (from HuES9.E1 cells-derived MSCs) have been shown to increase the formation of new alveolar bone in a rat model of surgically created periodontal intrabony defects. These EXOs could increase rat periodontal ligament cell migration and proliferation through CD73-mediated adenosine receptor activation of pro-survival Akt and ERK signaling pathways [93][97].