The potential, multifaceted therapeutic profile of cannabidiol (CBD), a major constituent derived from the Cannabis sativa plant, covers a wide range of neurological and psychiatric disorders, ranging from anxiety to pediatric epilepsy and drug addiction. However, the molecular targets responsible for these effects have been only partially identified. In this view, the involvement of the orexin system, the key regulator in arousal and the sleep/wake cycle, and in motivation and reward processes, including drug addiction, prompted us to explore, using computational and experimental approaches, the possibility that CBD could act as a ligand of orexin receptors, orexin 1 receptor of type 1 (OX1R) and type 2 (OX2R). Ligand-binding assays showed that CBD is a selective ligand of OX1R in the low micromolar range (Ki 1.58 ± 0.2 μM) while in vitro functional assays, carried out by intracellular calcium imaging and mobilization assays, showed that CBD acts as an antagonist at this receptor. Finally, the putative binding mode of CBD has been inferred by molecular docking and molecular dynamics simulations and its selectivity toward the OX1R subtype rationalized at the molecular level. This study provides the first evidence that CBD acts as an OX1R antagonist, supporting its potential use in addictive disorders and/or body weight regulation.
- orexin receptors
- cannabidiol
- molecular docking
- molecular dynamics
- calcium mobilization assay
1. Introduction
2. Discussion
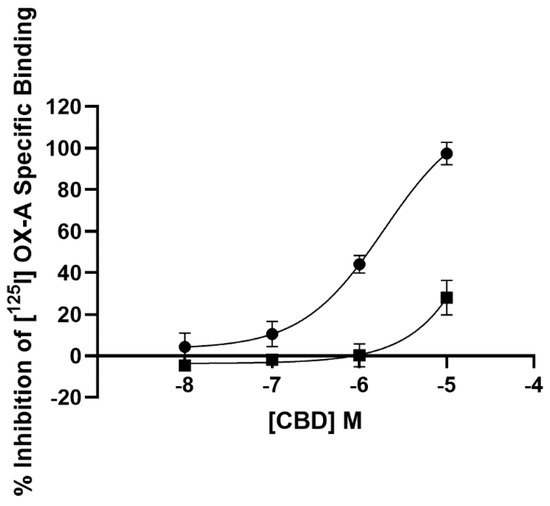
2.1. Radioligand Binding Assay
3.1. Radioligand Binding Assay
2.2. Calcium Mobilization Assay
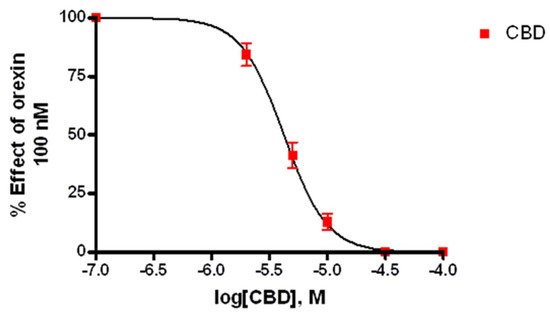
3.2. Calcium Mobilization Assay
23.3. Calcium Imaging
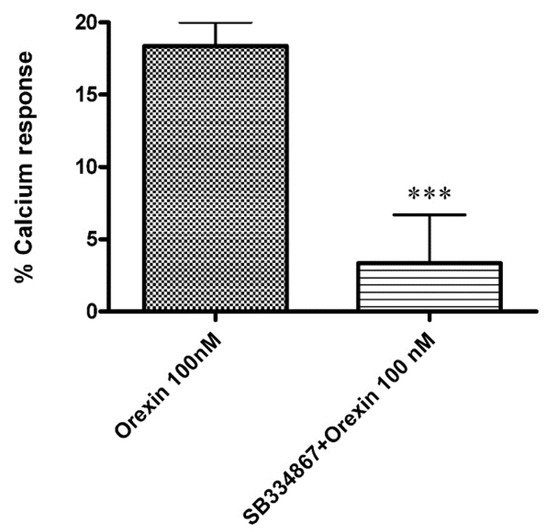
23.3.1. OX-A Increases Intracellular [Ca2+]i in CHO Cells Stably Expressing OX1R or OX2R
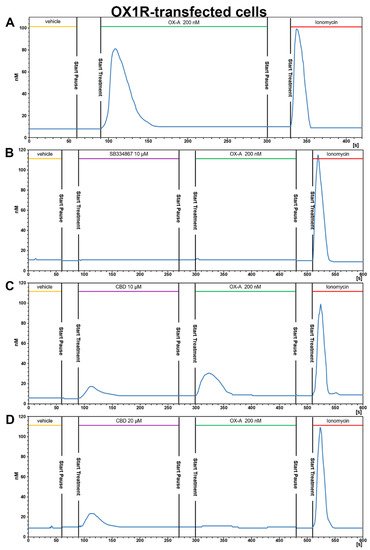
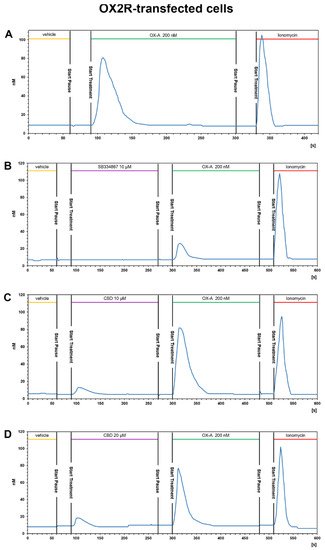
23.3.2. CBD Inhibits the Ca2+ Response Induced by OX-A in OX1R, but Not in OX2R Transfected CHO Cells
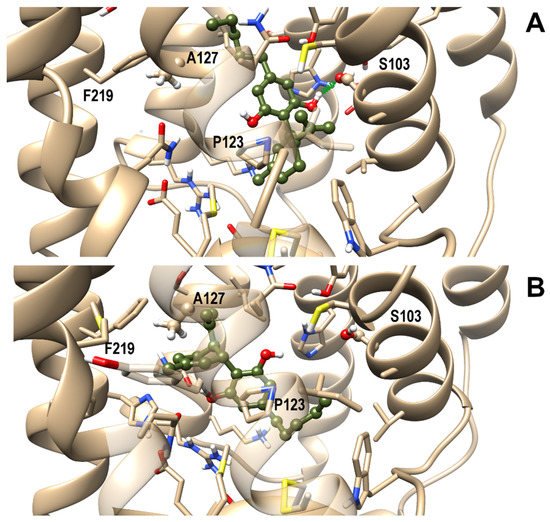
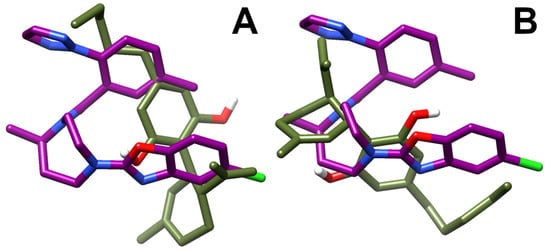
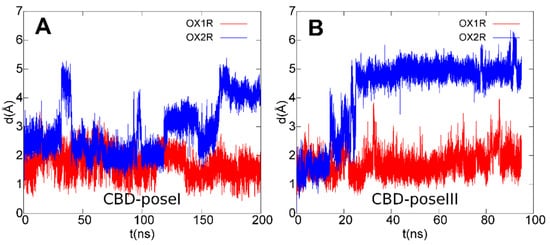
3.4. OX1R/CBD Theoretical Complex