Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Bo Li and Version 3 by Conner Chen.
Suture mesenchymal stem cells (SuSCs), a heterogeneous stem cell population, belong to mesenchymal stem cells (MSCs) or skeletal stem cells (SSCs), with the ability to self-renew and undergo multi-lineage differentiation. Unlike the well-established perivascular niche of SSCs in the long bone, stem cells of the cranial bone are generally located and confined within the cranial suture mesenchyme, subsequently defined as SuSCs. In the long bone, SSCs play an essential role in plenty of physiological processes, such as growth and development, life-long homeostasis, and fracture healing. Similarly, as the major stem cell population of cranial bones, the physiological significance of SuSCs is undoubted and self-evident.
- mesenchymal stem cells
- cranial sutures
- injury repair
1. The Anatomy of Cranial Sutures
The major sutures of the skull vault include the following: the metopic suture, also referred to as the frontal/interfrontal suture, extending from bregma to nasion, located between the two frontal bone plates; the sagittal suture extending from the bregma to lambda, located between the two parietal bone plates; the coronal suture extending from bregma to left/right pterion, located between the frontal bone plate and the left/right parietal bone plate; and the lambdoid suture extending from lambda to left/right asterion, located between the occipital bone plate and the left/right parietal bone plate (Figure 1).
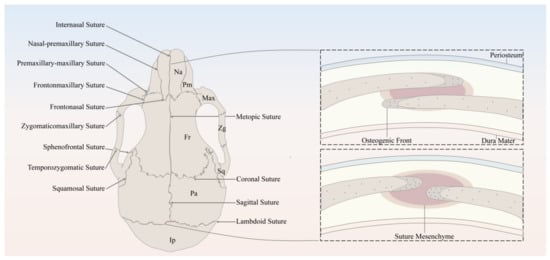
Figure 1. Schematic of the murine skull in the dorsal view, depicting the anatomy of cranial bones and sutures. Na, Nasal; Pm, Premaxilla; Max, Maxilla; Zg, Zygomatic; Sq, Squamosal; Fr, Frontal; Pa, Parietal; Ip, Interparietal.
It has been well established that the derivation of the skull vault comes from dual tissue lineages, namely, paraxial mesoderm and cranial neural crest [1][2][3][13,14,15]. As for the cranial sutures, not only do the sutures separate bones of different embryological origin, but they are themselves derived from different origins [3][15]. For instance, sutures derived from cranial neural crest include metopic sutures and sagittal sutures, while coronal sutures derived from paraxial mesoderm and the developmental origin of the lambdoid sutures remains unknown [1][3][13,15]. Differing in embryonic origin may lead to distinct capabilities of SuSCs in different sutures.
In spite of the discrepancy in anatomical locations and embryonic origins, cranial sutures have similarly fundamental features, which can be considered as a complex composed of four principal components [1][13], including the OFs of the approximating bone plates, the suture mesenchyme spanning the OFs, the overlying periosteum, and the underlying dura mater, which is also the outermost layer of meninges, namely, membranous coverings of the brain and spinal cord. To be noted, the OFs of the bone plates of the coronal and lambdoid sutures partially overlap each other, whereas the counterparts of the metopic and sagittal sutures abut end to end [1][13] (Figure 1).
Throughout the growth and development of the skull, the cranial suture mesenchyme remains unossified, which interposes between the OFs of the adjoining bone plates. The MSCs and osteoprogenitors residing along the OFs keep proliferating, subsequently differentiate into osteoblasts, and contribute to the new bone formation through intramembranous ossification [4][5][9,16], which happens with a direct differentiation into osteoblasts from MSCs and/or osteogenic precursors without assuming a chondrogenic fate. During the above-mentioned process, the osteoblasts will secrete a kind of extracellular collagen matrix called osteoid, which will then become mineralized and deposit at the leading edges of bone plates. As for the bone plates, they remain separated to allow the growth and expansion of the skull in concert with the growing brain [1][13].
2. Temporal and Spatial Distribution Features of SuSCs Subpopulations
As aforementioned, reporter constructs expressed by Gli1 [6][1], Axin2 [7][2], Prrx1 [8][3], and Ctsk [9][4] have been identified to label SuSCs specifically. These four SuSCs subpopulations are physically tightly related but do not fully overlap (Figure 2), reflecting the heterogeneity of MSCs to some degree. Therefore, the use of multiple markers together might delineate and define SuSCs better, on which to date no study has been done.
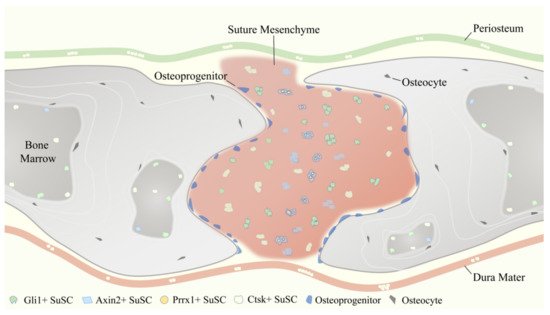
Figure 2. Temporospatial distribution pattern of four representative suture mesenchymal stem cells (SuSCs) subpopulations, including Gli1+ SuSCs, Axin2+ SuSCs, Prrx1+ SuSCs, and Ctsk+ SuSCs.
2.1. Temporal and Spatial Distribution Features of Gli1+ SuSCs
Gli1 is an essential transcription factor of the Hedgehog (Hh) signaling pathway. Currently, it has been widely validated as a common and reliable MSCs marker in a variety of tissues and organs throughout the whole body [10][39], including but not limited to the skull [6][1], long bones [11][12][40,41], incisors [13][14][42,43], periodontal ligament [15][44], intestine [16][45], and respiratory tract [17][46].
From P0 to 1 month after birth (P30) in mice, which is equivalent to the early stage of growth and development in humans, Gli1+ SuSCs gradually restrict the cranial suture from a widespread distribution pattern and are ultimately confined to the suture mesenchyme [6][1]. Specifically, Gli1+ SuSCs can be detected in the periosteum, dura mater, and suture mesenchyme from P0 to P14, whereas from P14 to P30, Gli1+ SuSCs are almost merely observed within the cranial suture region. In addition, Gli1+ SuSCs are also observable lining on the inner surface of the cranial bone marrow cavity, even though the number is neglectable. After 8 months of lineage tracing, Gli1+ SuSCs and their progenies are distributed throughout the calvarium, detectable in cranial suture mesenchyme, periosteum, and dura mater. Moreover, quite a few osteocytes are also labeled, indicating that they derive from Gli1+ SuSCs [6][1]. Additionally, our research group carried out short-term (30 days) and long-term (1 year) lineage tracing and verified the suture-specific distribution pattern of Gli1+ SuSCs independently. Recently, a tissue clearing method, the PEG associated solvent system (PEGASOS) [18][47], has been applied to render craniofacial bones entirely transparent and to delineate the temporospatial distribution of Gli1+ SuSCs with deep imaging techniques [19][48]. It has been convincingly demonstrated that Gli1+ SuSCs are spatially associated with vasculature during the postnatal craniofacial development [19][48], which implies Gli1+ SuSCs may bear a resemblance to MSCs of long bones, for instance, residing in a peri-vascular milieu.
2.2. Temporal and Spatial Distribution Features of Axin2+ SuSCs
Axin2 is a direct transcriptional target of β-catenin, as well as a negative regulator of the Wnt/β-catenin (Wnt) signaling pathway [20][30]. Unlike Gli1+ SuSCs, Axin2+ SuSCs do not exhibit a diffused distribution pattern in the calvarium in the early stages of postnatal growth and development and are rarely detected in either the periosteum or the dura mater. Starting from P0, Axin2+ SuSCs are primarily located in the cranial suture and concentrated in the midline of the suture mesenchyme. After 1-month, 3-month, and 1-year lineage tracing, Axin2+ SuSCs and their progeny cells continue to accumulate without showing any sign of diminishing. Besides cranial sutures, Axin2+ SuSCs and their derivatives are also widely expanded and found within periosteum and dura mater over a long tracing period; some of the cells are embedded in the bone plate as osteocytes. Even more than 1 year of tracing, the Axin2+ SuSCs and their derivatives remain detectable and keep increasing in all calvarial sutures except the posterior frontal suture, which generally undergoes fusion in juveniles [7][2].
2.3. Temporal and Spatial Distribution Features of Prrx1+ SuSCs
Prrx1 is a transcription factor highly expressed during limb bud formation [21][22][49,50], and craniofacial development [22][50]. In terms of localization, Prrx1+ SuSCs have their own unique characteristics. To illustrate, Prrx1+ SuSCs reside exclusively within the posterior frontal suture, coronal suture, sagittal suture, and lambdoid suture, but are absent in other craniofacial sutures, periosteum, and dura mater. Interestingly, the number of Prrx1+ SuSCs will decrease with age. The quantitative analysis of 8-, 16-, 24-, and 32-week-old mice showed that the total cellularity or the cell density of the suture did not change with age; however, the population of Prrx1+ SuSCs displayed a significant reduction with age continuously. The total number of Prrx1+ SuSCs in the coronal suture decreased up to 75% from 8 weeks of age to 32 weeks of age [8][3]. This result may attribute to the fact that the Prrx1-expressing cells contain a large proportion of osteoprogenitors or transit-amplifying cells (TACs), while the percentage of the bona fide SuSCs is low. Last but not least, Prrx1+ SuSCs can express Axin2, which will increase upon the stimulation of the Wnt agonist. Thereby, the researchers postulate that Prrx1+ SuSCs are a subset of Axin2+ SuSCs [8][3].
2.4. Temporal and Spatial Distribution Features of Ctsk+ SuSCs
For decades, Cathepsin K (Ctsk) has been widely conceived as a classic marker of osteoclasts in the field of bone research [23][51]. However, in the past few years, it has been revealed that Ctsk can label the mesenchymal progenitors in the perichondrial groove of Ranvier [24][52]. More recently, another group identified Ctsk-expressing stem cells located in the periosteum of long bone and cranial suture via the scRNA-seq approach, together with lineage tracing and a series of rigorous in vitro and in vivo experiments to verify their ‘stemness’ [9][4]. Leaving long bones alone and focusing solely on the skull, at P15 and P32, Ctsk+ SuSCs and their progenies were not only presented in the cranial sutures, but also in the overlying periosteum, underlying dura mater, and bone marrow cavity of the calvarium, which was indicated by the visible signals of membrane-bound green fluorescent protein (mGFP). Further, fluorescence-activated cell sorting (FACS) analysis of P6 calvarial tissues found that the percentage of Ctsk+ SuSCs in suture (31.1%) was significantly higher than the percentage of Ctsk+ SuSCs in calvarial periosteum (4.37%), suggesting the enrichment of Ctsk+ SuSCs in the suture region [9][4]. Therefore, cells with an immunophenotype of Ctsk+ SuSCs existed predominantly in the suture mesenchyme, which was also consistent with the notion that cranial sutures contain mesenchymal progenitors that migrated to the periosteum as maturation [6][7][1,2]. Although the majority of this study focused on exploring and discussing Ctsk+ periosteal stem cells (PSCs) in the long bone, it still provides some valuable insights on Ctsk+ SuSCs and their properties.
3. Characteristics of SuSCs and Their Role in Injury Repair
In general, the subpopulations of SuSCs labeled and distinguished by Gli1, Axin2, Prrx1, and Ctsk have similar but not identical biological characteristics. They all possess a self-renewal ability and multi-lineage differentiation potential (excluding Prrx1+ SuSCs, which were only tested for osteogenic differentiation [8][3]), and participate in calvarial bone injury repair (excluding Ctsk+ SuSCs, which did not have direct experimental evidence showing their involvement in calvaria injury repair, albeit Ctsk+ PSCs is proved to contribute to long bone fracture healing [9][4]).
3.1. Biological Characteristics of Different SuSCs Subpopulations
According to the International Society for Cellular Therapy (ISCT), a standard set of stem cell criteria must be fulfilled for defining MSCs [25][26][53,54], including plastic-adherent ability when cultured in vitro, specific surface markers expression (such as CD73, CD90, and CD105), and trilineage (osteogenic, chondrogenic, and adipogenic) differentiation potential. Of note, this definition is based on cultured cells, and it remains largely unknown regarding the criteria applied to identify MSCs in vivo. Hence, several crucial biological characteristics of each subset of SuSCs are reviewed here accordingly, including self-renewal ability, multipotency, and the expression of various markers of MSCs or SSCs.
As for the capacity to self-renew, both Gli1+ SuSCs and Prrx1+ SuSCs are tested through long-term lineage tracing, which somehow reflects the self-renewal ability of labeled cells, but have not been discussed in detail [6][8][1,3]. Axin2+ SuSCs have gone through long-term lineage tracing without decreasing in number; meanwhile, EdU (5-ethynyl-2′-deoxyuridine) assay and Ki67 immunofluorescence pointed out that Axin2+ SuSCs were slow cycling in nature instead of active proliferating, indicated by their label-retaining property [7][2]. For Ctsk+ SuSCs, mesensphere assays were performed consecutively for three rounds, which aimed to critically evaluate the self-renewal ability in vitro. As a result, more than 60% of Ctsk+ SuSCs were able to form primary and secondary mesenspheres, whereas tertiary mesensphere formation was significantly reduced [9][4].
Concerning the potential of multi-lineage differentiation, Gli1+ SuSCs were capable of trilineage differentiation under induction in vitro [6][1]. When comparing the differentiation potential of Gli1+ SuSCs with bone marrow-derived MSCs (BMMSCs) from the same mice, researchers noticed that Gli1+ SuSCs were more inclined to differentiate into osteoblasts and chondrocytes because of their remarkably weaker adipogenic performance [6][1]. For Axin2+ SuSCs, kidney capsule transplantation studies demonstrated that Axin2+ SuSCs could form ectopic bone in vivo without any intervention and therefore possessed the osteogenic ability. In addition, the transplanted Axin2+ SuSCs were able to generate cartilage with the presence of bone morphogenetic protein 2 (BMP2), suggesting their chondrogenic potential [7][2]. Unlike Axin2+ SuSCs, Ctsk+ SuSCs could only undergo osteogenesis in vivo, which was revealed by the kidney capsule transplantation experiment, and could not give rise to cartilage, because Ctsk+ SuSCs were intramembranous-competent and predominantly orchestrated the process of intramembranous ossification. However, Ctsk+ SuSCs displayed clonal multipotency in vitro for differentiation into osteoblasts, chondrocytes, and adipocytes [9][4]. Prrx1+ SuSCs were only tested and verified for their osteogenic differentiation ability upon appropriate induction (recombinant mouse WNT3a) [8][3].
Regarding the expression of MSC or SSC markers, FACS analysis revealed that Gli1+ SuSCs and their derivatives highly expressed typical MSC markers, including CD44, CD73, CD90, Sca1, and CD146, but did not express CD34 [6][1]. Astonishingly, immunohistochemical staining showed that most Gli1+ SuSCs did not express CD44, CD73, CD90, Sca1, and CD146 in vivo. By microarray and real-time RT-PCR, it was found that Axin2+ SuSCs highly expressed Leptin receptor (LepR) and Gli1, while the expression levels of Nestin, Gremlin1, and CD146 were similar to Axin2− suture cell populations [7][2]. Similarly, it was detected by real-time RT-PCR that Prrx1+ SuSCs exhibited an elevation of Pdgfrα and CD146 compared with Prrx1− cells isolated from calvarial sutures [8][3]. As for Ctsk+ SuSCs, CD45− Ter119− CD31− (Lin−) non-hematopoietic and non-vascular endothelial Ctsk-mGFP cells [27][28][55,56] were initially sorted by FACS, and subsequently, the expression of SSC markers was examined in the sorted Ctsk-mGFP cells. Finally, Thy1.2−, 6C3−, CD49flow, CD51low, CD200+, CD105− Ctsk-mGFP cells were determined as Ctsk+ SuSCs, which did not express CD146 and Sca1 [9][29][4,57].
3.2. Contribution of SuSCs in Calvarial Bone Injury
SuSCs play an indispensable role in the injury repair and tissue regeneration of calvarial bone defects after birth [30][58]. Studies have shown that Gli1+ SuSCs were rapidly activated into proliferation within 24 h after an injury occurs. Two weeks after experimental injury, most of the infiltrated cells within the injury site were labeled, indicating their derivation from Gli1+ SuSCs. One month after experimental injury, the periosteum, dura mater, and osteocytes in the re-ossified region were labeled, suggesting that Gli1+ SuSCs contribute to calvarial bone defect repair [6][1]. Additionally, Gli1-CreERT2;R26-ZsGreenflox mice were induced and their calvaria (skull bone flaps containing the sagittal suture) were dissected under sterile conditions and transplanted into nude mice, which were used as the recipient mice with a calvarial window for placing transplants. It was found that the suture transplants integrated into the host bone and healing were achieved one-month post-surgery, with a significant number of cells within the periosteum, dura mater, and bone of the transplant strongly labeled. On the contrary, transplants not containing any suture tissue (with periosteum and dura mater preserved) from the same donor mice were transplanted and served as controls and ended up with poor healing and failure in generating new periosteum, dura mater, or osteocytes [6][1]. Thus, the cranial sutures and the resident Gli1+ SuSCs are the main sources of reparative cells functioning in calvarial bone injury repair; the periosteum and dura mater are either unable or insufficient to accomplish efficient calvarial bone regeneration. Similarly, Axin2+ SuSCs also respond to calvarial bone injury and promptly expand within the suture mesenchyme. Four weeks after experimental injury, a drastic expansion of Axin2+ SuSCs has been observed surrounding the skeletogenic suture mesenchyme. Further, Axin2+ SuSCs moved into the injury site and co-localized with Osx+ osteoprogenitors and Sost+ osteocyte, indicating their direct contribution to the injury repair of the skull. When Axin2+ SuSCs were isolated from Axin2Cre-Dox;R26RlacZ mice and directly implanted into the injury site, enhancements of the healing process were detected at two and four weeks after the operation. In comparison, neither transferring Axin2− cells nor implanting without any cells serving as control did not show significant improvement [7][2]. As expected, 5 days, 10 days, and 30 days after experimental injury, Prrx1+ SuSCs and their progenies were found to contribute to the repair and regeneration of neural crest-derived (frontal) and mesoderm-derived (parietal) calvarial bones [8][3]. Plus, the parietal bone defects were unable to heal if the surrounding coronal and sagittal sutures were surgically removed concomitantly to the creation of the defect, while removal of the sutures away from the parietal bone defects did not affect the healing process [8][3]. Regarding Ctsk+ SuSCs, it has not been tested through the calvarial bone defect model to evaluate their performance in injury repair. However, based on the pivotal role of Ctsk+ PSCs in the process of long bone fracture healing [9][4], we speculate that Ctsk+ SuSCs should facilitate calvarial bone healing as well.