The myotendinous junction (MTJ) is the muscle-tendon interface and constitutes an integrated mechanical unit to force transmission. Joint immobilization promotes muscle atrophy via disuse, while physical exercise can be used as an adaptative stimulus. In this study, we aimed to investigate the components of the MTJ and their adaptations and the associated elements triggered with aquatic training after joint immobilization. Forty-four male Wistar rats were divided into sedentary (SD), aquatic training (AT), immobilization (IM), and immobilization/aquatic training (IMAT) groups. The samples were processed to measure fiber area, nuclear fractal dimension, MTJ nuclear density, identification of telocytes, sarcomeres, and MTJ perimeter length. In the AT group, the maintenance of ultrastructure and elements in the MTJ region were observed; the IM group presented muscle atrophy effects with reduced MTJ perimeter; the IMAT group demonstrated that aquatic training after joint immobilization promotes benefits in the muscle fiber area and fractal dimension, in the MTJ region shows longer sarcomeres and MTJ perimeter. We identified the presence of telocytes in the MTJ region in all experimental groups. We concluded that aquatic training is an effective rehabilitation method after joint immobilization due to reduced muscle atrophy and regeneration effects on MTJ in rats.
- nuclear domain
- sarcomere
- telocyte
- muscle-tendon perimeter
- aquatic training
- joint immobilization
1. Introduction
2.Aquatic Training after Joint Immobilization in Rats Promotes Adaptations in Myotendinous Junctions
2.1. Fiber Area
The AT group had a lower fiber area than the SD group (p = 0.0001). The IM group had a lower fiber area than the SD (p < 0.0001) and AT groups (p < 0.0001). By contrast, the IMAT group had a lower fiber area than the SD group (p = 0.0001) and a higher fiber area than the IM group (p < 0.0001) (Figure 1).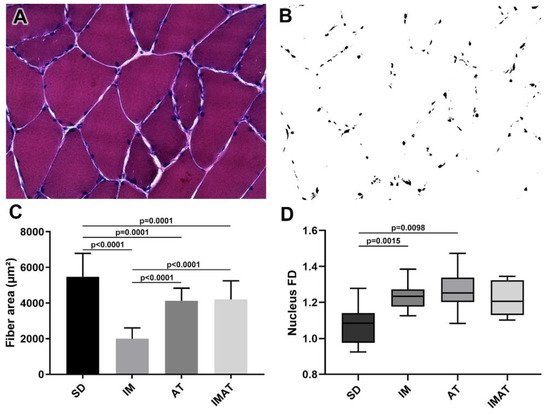
2.2. Fractal Dimension (FD)
The AT (p = 0.0015) and IM (p = 0.0098) groups had a higher FD than the SD group (Figure 1).2.3. MTJ Nuclear Density
The IM group had a lower MTJ nuclear density than the AT (p = 0.0002) and IMAT (p = 0.0005) groups. The other groups showed no significant difference (Figure 2).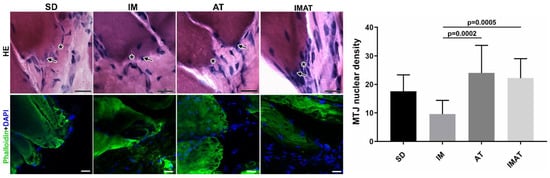
2.4. MTJ Morphology
Through the ultrastructural descriptions of the MTJ, its morphology and plasticity as well as associated elements were revealed, i.e., telocytes (Figure 3). We observed branched sarcoplasmatic invaginations and a telocyte surrounding the MTJ near a blood capillary in the SD group.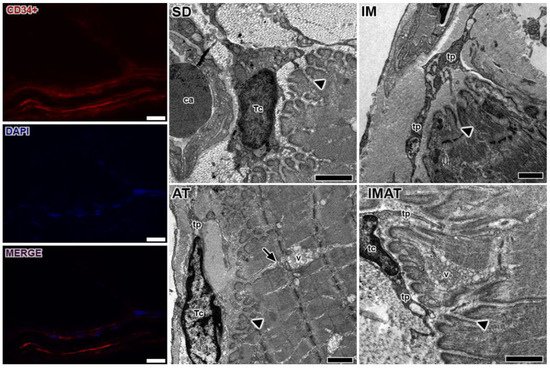
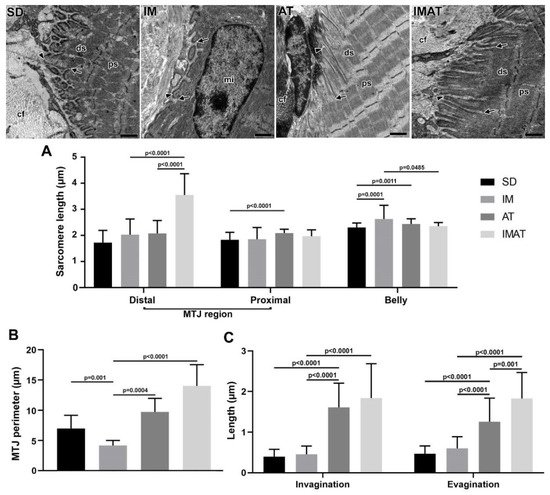
2.5. Sarcomeres
The distal sarcomere had longer lengths in the IMAT group than in the IM (p < 0.0001) and AT groups (p < 0.0001). The AT group had longer proximal than the SD group (p < 0.0001). The belly sarcomeres were longer in the IM group than in the SD (p = 0.0001) and IMAT groups (p = 0.0485); the AT group had longer sarcomeres than the SD group (p = 0.0011) (Figure 4).2.6. MTJ Morphometry
The IM group demonstrated a shorter MTJ perimeter than the SD (p = 0.001) and AT (p = 0.0004) groups. By contrast, the lengths of sarcoplasmatic invagination and evagination in the IM group were not significantly different from the SD group (p > 0.05) but were lower than the AT (p < 0.0001) and IMAT (p < 0.0001) groups. The AT group showed a higher MTJ perimeter than the IM group (p = 0.0004). Moreover, the AT group had longer sarcoplasmatic invagination and evagination lengths than the SD (p < 0.0001) and AT groups (p < 0.0001). The IMAT group had longer MTJ perimeter (p < 0.0001) and longer sarcoplasmatic (p < 0.0001) invaginations than the IM group. In addition, the IMAT group had longer sarcoplasmatic evaginations than the IM (p < 0.0001) and AT groups (p = 0.001) (Figure 4).3. Discussion
The results of this study revealed the joint immobilization effect of muscle atrophy due to disuse. The study also showed the effect and repercussion of AT as a rehabilitation method for the muscle belly and the muscle-tendon complex as well as other parameters, including the nucleus, sarcomere, contact perimeter, and telocyte.References
- Ciena, A.P.; Luques, I.U.; Dias, F.J.; Yokomizo de Almeida, S.R.; Iyomasa, M.M.; Watanabe, I.S. Ultrastructure of the myotendinous junction of the medial pterygoid muscle of adult and aged Wistar rats. Micron 2010, 41, 1011–1014.
- Natali, L.H.; da Silva, T.S.; Ciena, A.P.; Padoin, M.J.; Alves, É.P.B.; Aragão, F.A.; Bertolini, G.R.F. Efeitos da corrida em esteira em músculos sóleos de ratos encurtados por imobilização. Rev. Bras. Med. Do Esporte 2008, 14, 490–493.
- Kannus, P.; Jozsa, L.; Kvist, M.; Lehto, M.; Jarvinen, M. The effect of immobilization on myotendinous junction: An ultrastructural, histochemical and immunohistochemical study. Acta Physiol. Scand. 1992, 144, 387–394.
- Curzi, D. Ultrastructural study of myotendinous junction plasticity: From disuse to exercise. Sport Sci. Health 2016, 12, 279–286.
- Perillo, M.; Folker, E.S. Specialized Positioning of Myonuclei Near Cell-Cell Junctions. Front. Physiol. 2018, 9, 1–10.
- Pimentel Neto, J.; Rocha, L.C.; Barbosa, G.K.; Jacob, S.; Krause Neto, W.; Watanabe, I.; Ciena, A.P. Myotendinous junction adaptations to ladder-based resistance training: Identification of a new telocyte niche. Sci. Rep. 2020, 10, 1–8.
- Popescu, L. Telocytes-a novel type of interstitial cells. Recent Res. Mod. Med. 2011, 11, 424–432.
- Edelstein, L.; Fuxe, K.; Levin, M.; Popescu, B.O.; Smythies, J. Telocytes in their context with other intercellular communication agents. Semin. Cell Dev. Biol. 2016, 55, 9–13.
- Marini, M.; Rosa, I.; Ibba-Manneschi, L.; Manetti, M. Telocytes in skeletal, cardiac and smooth muscle interstitium: Morphological and functional aspects. Histol. Histopathol. 2018, 33, 1151–1165.
- Rocha, L.C.; Jacob, C.d.S.; Barbosa, G.K.; Neto, J.P.; Neto, W.K.; Gama, E.F.; Ciena, A.P. Remodeling of the skeletal muscle and postsynaptic component after short-term joint immobilization and aquatic training. Histochem. Cell Biol. 2020, 154, 621–628.
- Cação-Benedini, L.O.; Ribeiro, P.G.; Prado, C.M.; Chesca, D.L.; Mattiello-Sverzut, A.C. Immobilization and therapeutic passive stretching generate thickening and increase the expression of laminin and dystrophin in skeletal muscle. Braz. J. Med. Biol. Res. 2014, 47, 483–491.
- Carvalho, L.C.; Polizello, J.C.; Padula, N.; Freitas, F.C.; Shimano, A.C.; Mattiello-sverzut, A.C. Mechanical properties of gastrocnemius electro stimulated after immobilization. Acta Ortop Bras 2009, 17, 269–272.
- Cação-Benedini, L.O.; Ribeiro, P.G.; Gomes, A.R.S.; Ywazaki, J.L.; Monte-Raso, V.V.; Prado, C.M.; Mattiello-Sverzut, A.C. Remobilization through stretching improves gait recovery in the rat. Acta Histochem. 2013, 115, 460–469.