Drought and salinity are major constraints to agriculture. Here, we present an overview of the global situation and the consequences of drought and salt stress connected to climatic changes.
- osmotic adjustment
- ionic
- redox homeostasis
- transcription factors
- salt-responsive genes
- genetic diversity
- germplasm
- cross-transferability
1. Global Situation
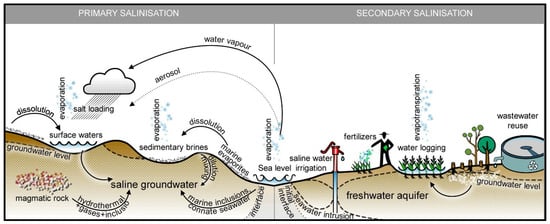
2. Germplasm
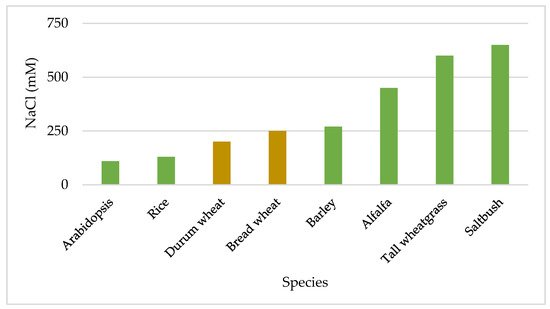
3. Morphological and Physiological Response
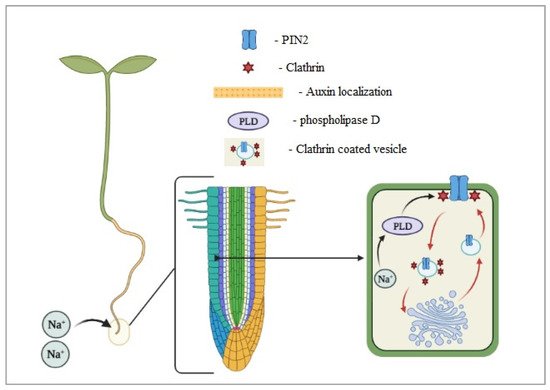
4. Mechanisms of Tolerance
Osmotic Stress | Ionic Stress | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
Process | Candidate Genes | Osmotic Tolerance | Na+ Excluding | Tissue Tolerance | References | |||||||||||||||||||
Signaling | SOS3, | SnRKs | Signaling regulation | Activation of ion antiporter | Regulation of vacuolar loading |
[77] |
[30] |
|||||||||||||||||
Photosynthesis | ERA1, | PP2C , AAPK, PKS3 | Stomatal closure | regulation | Protection of chloroplast | from ion toxicity | Delay Na+ toxicity | effect in chloroplast | ||||||||||||||||
Accumulation of Na+ | in shoots | HKT, SOS1 | - | Decreasing long | distance transport of Na + | Decreasing energy used on Na+ exclusion | ||||||||||||||||||
Accumulation of Na+in vacuoles | NHX, AVP | - | Increased sequestration of Na+ | into root vacuoles | Increased sequestration of Na+ | into leaf vacuoles | ||||||||||||||||||
Accumulation | of organic solutes | P5CS, OTS, MT1D, M6PR, S6PDH, IMT1 | Increasing osmotic adjustment | Reduction of Na+ accumulation | Accumulation of organic solutes in cytoplasm | |||||||||||||||||||
References
- Mann, M.E.; Gleick, P.H. Climate change and California drought in the 21st century. Proc. Natl. Acad. Sci. USA 2015, 112, 3858–3859.
- Nam, W.H.; Hayes, M.J.; Svoboda, M.D.; Tadesse, T.; Wilhite, D.A. Drought hazard assessment in the context of climate change for South Korea. Agric. Water Manag. 2015, 160, 106–117.
- Bot, A.; Benites, J. Drought-resistant soils: Optimization of Soil Moisture for Sustainable Plant Production. In Proceedings of the Electronic Conference ‘Drought-Resistant Soils. Optimization of Soil Moisture for Sustainable Plant Production.’ FAO Land and Water Development Division, 15 November–18 December 2004; Volume 11.
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S.; et al. Climate change and interconnected risks to sustainable development in the Mediterranean. Nat. Clim. Chang. 2018, 8, 972–980.
- Lawlor, D.W.; Mitchell, R.A. Crop ecosystem responses to climatic change. In Climate Change and Global Crop Productivity; Nobel, P.S., Reddy, K.R., Hodges, H.F., Eds.; CABI Publishing: Wallingford, UK, 2001; pp. 57–80.
- Huang, J.; Zhang, W.; Zuo, J.; Bi, J.; Shi, J.; Wang, X.; Chang, Z.; Huang, Z.; Yang, S.; Zhang, B.; et al. An overview of the semi-arid climate and environment research observatory over the loess plateau. Adv. Atmos. Sci. 2008, 25, 906–921.
- Houghton, J.T.; Albritton, D.L.; Meira Filho, L.G.; Cubasch, U.; Dai, X.; Ding, Y.; Karl, T. Technical summary of working group 1. In Intergovernmental Panel on Climate Change (IPCC), Climate Change 2001: The Scientific Basis; Cambridge University Press: Cambridge, UK, 2001; pp. 3–45.
- Rani, G.; Kaur, J.; Kumar, A.; Yogalakshmi, K.N. Ecosystem health and dynamics: An indicator of global climate change. In Contemporary Environmental Issues and Challenges in Era of Climate Change; Springer: Singapore, 2019; pp. 1–32.
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131.
- Wegulo, S.; Giesler, L.; Harveson, R.; Jackson-Ziems, T.; Liu, B.; Korus, K. 2013 Crop Production Clinic Proceedings, Impacts of Drought on Disease Development and Management. Pap. Plant Pathol. 2013, 125–127.
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 153–188.
- Daliakopoulos, I.N.; Tsanis, I.K.; Koutroulis, A.; Kourgialas, N.N.; Varouchakis, A.E.; Karatzas, G.P.; Ritsema, C.J. The threat of soil salinity: A European scale review. Sci. Total Environ. 2016, 573, 727–739.
- Allen, M.R.; Ingram, W.J. Constraints on future changes in climate and the hydrologic cycle. Nature 2002, 419, 228–232.
- Rozema, J.; Flowers, T.J. Crops for salinized world. Science 2008, 322, 1478–1480.
- Konapala, G.; Mishra, A.K.; Wada, Y.; Mann, M.E. Climate change will affect global water availability through compounding changes in seasonal precipitation and evaporation. Nat. Commun. 2020, 11, 1–10.
- Ali, S.; Rizwan, M.; Qayyum, M.F.; Ok, Y.S.; Ibrahim, M.; Riaz, M.; Arif, M.S.; Hafeez, F.; Al-Wabel, M.I.; Shahzad, A.N. Biochar soil amendment on alleviation of drought and salt stress in plants: A critical review. Environ. Sci. Pollut. Res. 2017, 24, 12700–12712.
- Ahmad, P.; Prasad, M.N.V. Abiotic Stress Responses in Plants: Metabolism, Productivity and Sustainability; Springer Science + Business Media, LLC: New York, NY, USA, 2012.
- Lawlor, D.W. Stress metabolism: Its implication in breeding programmes. In Drought Tolerance in Winter Cereals; Srivastava, E.J.P., Porceddu, E., Acevedo, S., Eds.; Varma. Icarda: Aleppo, Syria, 1987.
- Hatfield, J.L.; Dold, C. Water-use efficiency: Advances and challenges in a changing climate. Front. Plant Sci. 2019, 10, 103.
- Richards, R.A.; Passioura, J.B. Seminal Root Morphology and Water Use of Wheat I. Environmental Effects 1. Crop Sci. 1981, 21, 249–252.
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681.
- Shavrukov, Y.; Kurishbayev, A.; Jatayev, S.; Shvidchenko, V.; Zotova, L.; Koekemoer, F.; De Groot, S.; Soole, K.; Langridge, P. Early flowering as a drought escape mechanism in plants: How can it aid wheat production? Front. Plant Sci. 2017, 8, 1950.
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. In Sustainable Agriculture; Springer: Cham, Switzerland, 2009; pp. 153–188. ISBN 9789048126651.
- Nieves-Cordones, M.; García-Sánchez, F.; Pérez-Pérez, J.G.; Colmenero-Flores, J.M.; Rubio, F.; Rosales, M.A. Coping With Water Shortage: An Update on the Role of K+, Cl-, and Water Membrane Transport Mechanisms on Drought Resistance. Front. Plant Sci. 2019, 10, 1619.
- Galvan-Ampudia, C.S.; Julkowska, M.M.; Darwish, E.; Gandullo, J.; Korver, R.A.; Brunoud, G.; Haring, M.A.; Munnik, T.; Vernoux, T.; Testerink, C. Halotropism is a response of plant roots to avoid a saline environment. Curr. Biol. 2013, 23, 2044–2050.
- Dai, M.; Zhang, C.; Kania, U.; Chen, F.; Xue, Q.; McCray, T.; Li, G.; Qin, G.; Wakeley, M.; Terzaghi, W.; et al. A PP6-type phosphatase holoenzyme directly regulates PIN phosphorylation and auxin efflux in Arabidopsis. Plant Cell 2012, 24, 2497–2514.
- Sun, F.; Zhang, W.; Hu, H.; Li, B.; Wang, Y.; Zhao, Y.; Li, K.; Liu, M.; Li, X. Salt modulates gravity signaling pathway to regulate growth direction of primary roots in arabidopsis. Plant Physiol. 2008, 146, 178–188.
- Julkowska, M.M.; Koevoets, I.T.; Mol, S.; Hoefsloot, H.; Feron, R.; Tester, M.A.; Keurentjes, J.J.B.; Korte, A.; Haring, M.A.; De Boer, G.J.; et al. Genetic components of root architecture remodeling in response to salt stress. Plant Cell 2017, 29, 3198–3213.
- Mondini, L.; Pagnotta, M.A. Drought and Salt Stress in Cereals; Springer: Cham, Switzerland, 2015; pp. 1–31.
- Sun, T.; Wang, Y.; Wang, M.; Li, T.; Zhou, Y.; Wang, X.; Wei, S.; He, G.; Yang, G. Identification and comprehensive analyses of the CBL and CIPK gene families in wheat (Triticum aestivum L.). BMC Plant Biol. 2015, 15, 1–17.
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. CRC. Crit. Rev. Plant Sci. 2005, 24, 23–58.
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273.
- Faïc Xal Brini, F.; Hanin, M.; Mezghani, I.; Berkowitz, G.A.; Masmoudi, K. Overexpression of wheat Na+/H+ antiporter TNHX1 and H+-pyrophosphatase TVP1 improve salt-and drought-stress tolerance in Arabidopsis thaliana plants. J. Exp. Bot. 2007, 58, 301–308.
- Ramezani, A.; Niazi, A.; Abolimoghadam, A.A.; Zamani Babgohari, M.; Deihimi, T.; Ebrahimi, M.; Akhtardanesh, H.; Ebrahimie, E. Quantitative expression analysis of TaSOS1 and TaSOS4 genes in cultivated and wild wheat plants under salt stress. Mol. Biotechnol. 2013, 53, 189–197.
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663.
- Zhang, Y.M.; Zhang, H.M.; Liu, Z.H.; Li, H.C.; Guo, X.L.; Li, G.L. The wheat NHX antiporter gene TaNHX2 confers salt tolerance in transgenic alfalfa by increasing the retention capacity of intracellular potassium. Plant Mol. Biol. 2015, 87, 317–327.
- Tavakoli, M.; Poustini, K.; Alizadeh, H. Proline accumulation and related genes in wheat leaves under salinity stress. J. Agric. Sci. Technol. 2016, 18, 707–716.