2. Structural Organization of the Shell
Encapsulin shells are icosahedral (12 vertices, 20 faces, 30 edges) complexes formed by self-assembly of protomer proteins
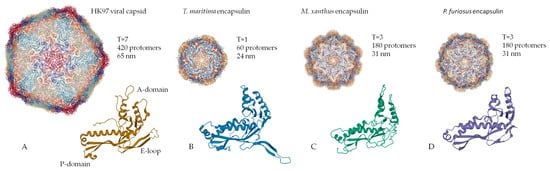
[2][13][24][2,13,24]. Encapsulin shell proteins are homologous to gp5-HK97 phage main capsid protein (A), as evidenced by the structural similarity between the gp5 protomer protein and encapsulin shell protein
[26]. It should be noted that the HK97 fold is widespread in nature and has been observed in all other tailed phages
[27], in herpesvirus capsids
[28], and in the archaevirus HSTV-1
[29]. Like gp5, the encapsulin shell proteins oligomerize to form a complete nanocompartment. As well as viral capsids, encapsulin shell proteins can self-assemble into icosahedrons of different sizes. For example, encapsulins from
Pyrococcus furiosus and
Myxococcus xanthus consist of 180 protomers (30‒32 nm in diameter) (C,D), while those from
Thermotoga maritima are composed of 60 protomers (24 nm in diameter) (B) and encapsulins from
Quasibacillus thermotolerans consist of 240 protomers (42 nm in diameter)
[2]. To classify this structural organization, a triangulation number can be used. The triangulation number (T) is a virology term describing the icosahedral packaging of viral capsid structural elements and is the quotient of dividing the number of subunits in a capsid by 60
[30]. Additionally, T can be explained as the number of subdivisions of each triangular facet of icosahedrons into identical equilateral triangles. The number of these triangles is equal to T. Following this classification, the
Pyrococcus furiosus and
Myxococcus xanthus encapsulins form icosahedrons with T equal to 3,
Quasibacillus thermotolerans forms T = 4 icosahedrons, and
Thermotoga maritima forms T = 1 icosahedron. The structure of an encapsulin shell protomer protein, like HK97 phage gp5 protomer, has three conserved domains: a peripheral domain (P), an axial domain (A), and an elongated loop (E)
[2][6][14][30][31][2,6,14,30,31]. While the
Pyrococcus furiosus and
Myxococcus xanthus encapsulin protomer structures match the HK97 gp5 protomer structure well
[13][24][13,24], only the A and E domains from the
Thermotoga maritima encapsulin align well with those of the
Pyrococcus furiosus and
Myxococcus xanthus encapsulin protomer structures
[2]. In particular, the E loop from
Thermotoga maritima is shorter and rotated relative to the E loops of HK97 phage,
Pyrococcus furiosus, and
Myxococcus xanthus [2]. This allows the E loop to form a beta sheet with the E loop of a neighboring protomer, creating a tight interaction
[2]. This may explain why the
Thermotoga maritima encapsulin forms a T = 1 capsid, while the
Pyrococcus furiosus and
Myxococcus xanthus nanocompartments can form a larger T = 3 capsid.
Figure 1. Structural comparison of the capsomers/monomers and assembled capsid/encapsulins: (A) HK97 phage, (B) Thermotoga maritima encapsulin, (C) Myxococcus xanthus encapsulin, (D) Pyrococcus furiosus encapsulin. (PDB-ID: 2FT1, 3DKT, 4PT2, and 2E0Z, respectively).
It has been proposed that encapsulins and certain phage capsids have a common evolutionary origin
[2][32][2,32]. This hypothesis is supported by the identification of genes encoding phage-like proteins close to the encapsulin gene in the archaeon
Sulfolobus solfataricus [33].
Nanocompartments are also known to have multiple pores formed at the protomer junctions; the pore diameter is about 5 Å
[2][13][2,13]. These holes are likely to serve as a permeability barrier for larger molecules while allowing small molecules and ions to pass across the shell. There is evidence that substrates of encapsulated enzymes, such as hydrogen peroxide or ferrous iron, can cross the shell, while proteins or other large molecules, such as DNA or polysaccharides, are not able to pass
[17][18][22][25][17,18,22,25]. Interestingly, Williams et al. achieved an enlargement of pore diameter in the
Thermotoga maritima encapsulin shell up to 11 Å using site-directed mutagenesis
[34].
3. Cargo Proteins
The function of the nanocompartment is associated with the function of its protein cargo. The first insight into the mechanism of cargo encapsulation was obtained when the crystal structure of the
Thermotoga maritima encapsulin was investigated in detail. X-ray crystallography revealed a small amount of extra electron density related to a hydrophobic pocket on the luminal surface of the encapsulin shell corresponding to a short (about 10 amino acid) C-terminal sequence of a ferritin-like protein (FLP), found next to the encapsulin shell gene in the
Thermotoga maritima genome
[2]. Bioinformatic analysis revealed that this C-terminal sequence is conserved in different species where the cargo protein genes and encapsulin shell genes are located together in a putative operon. Examples of such “predicted” cargo proteins include: FLP, DyP (dye-decolorizing peroxidase), hemerythrin, and rubrerythrin
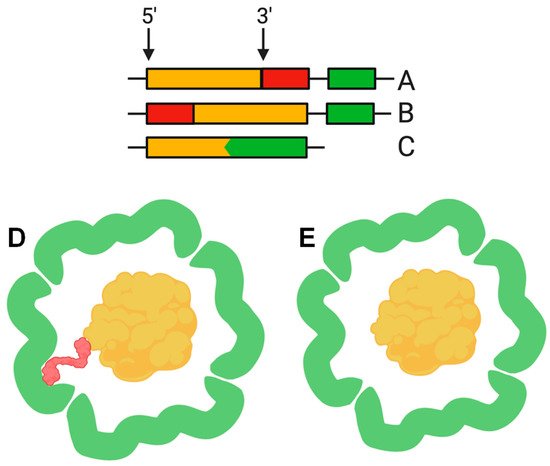
[2][20][2,20]. This C-terminal sequence, hereinafter termed the cargo loading peptide (CLP), is demonstrated to be sufficient for the loading of a cargo protein into the nanocompartment shell. The CLP coding sequence can be located at the 3′ or 5′ end of the cargo protein encoding gene (A,B,D). Deletion of CLP from the cargo protein disrupts encapsulation, while fusion of the CLP with the C-terminus of heterologous proteins, such as green fluorescent protein or luciferase, is enough for loading
[2][15][17][18][20][23][2,15,17,18,20,23]. Nevertheless, there are alternative models describing the interaction between shell proteins and cargo proteins. In some cases, such as the
Pyrococcus furiosus encapsulins, CLP is absent but the gene encoding the shell is fused with the gene encoding the cargo protein, forming a single polypeptide (C,E)
[13]. In the encapsulins found in
Firmicute bacteria, loading may occur partially via an N-terminus of CLP. Such a nanocompartment contains both an iron-mineralizing encapsulin-associated firmicute protein (IMEF) with C-terminal CLP and a ferredoxin protein with an N-terminal CLP
[6].
Figure 2. The cargo loading peptide coding sequence can be located at either the 5′ or 3′ end of the cargo loading peptide gene (A,B) with formation of the encapsulin structure (D). In some cases, the cargo protein encoding gene and encapsulin shell encoding gene are fused (C), and there is no need for a cargo-loading peptide to form encapsulin structure (E).
There is evidence that several cargo proteins can be loaded into a single encapsulin shell. For example, three different cargo proteins (EncB, EncC, and EncD) were detected in
Myxococcus xanthus encapsulins
[24], while in
Mycobacterium tuberculosis the following proteins were found: Mt-DyP, Mt-BfrB (bacterioferritin), and Mt-FolB (folate biosynthesis enzyme)
[23]. Another important issue related to loading is the fraction of the cargo protein within the nanocompartment. Each shell protein in encapsulin has a CLP binding site, but the amount of cargo is obviously limited by the shell volume. Modeling studies have shown that it is impossible to achieve a cargo-to-protein protomer ratio of more than 1:1 due to steric hindrance
[2]. An example is the
Brevibacterium linens nanocompartment with the mentioned DyP enzyme as a cargo protein that assembles into a hexamer (trimer of dimers) 89 Å in diameter. Steric hindrances were predicted to limit the loading to one hexamer per nanocompartment
[2], and measurements by native mass spectrometry confirmed the presence of six DyP monomers in the T = 1 nanocompartment, resulting in a cargo-to-protein protomer ratio of 1:10
[35].
4. Nanocompartments and Nanotechnology
Over the last decade, a variety of nanoscale targeted delivery systems based on micelles
[36][37][47,48], liposomes
[38][49], inorganic
[39][50] and polymer
[40][51] nanoparticles, and protein compartments
[41][42][52,53] have been developed. The small size and easy surface functionalization of these particles can increase the efficiency of drug delivery and accumulation compared with conventional nonencapsulated drugs. Such drugs are characterized by improved tissue penetration, longer circulation in the bloodstream, and reduced side effects
[43][54]. Additionally, targeted drug delivery systems can play a significant role in the diagnosis of diseases by interacting with specific molecular markers expressed in a particular pathology. However, none of the existing nanoscale systems (liposomes, inorganic and polymer nanoparticles) can be encoded into the genome and self-assembled by the cell, whereas the use of encapsulins opens up prospects for this particular application. Encapsulins are mainly used as a diagnostic tool or as a drug delivery system.
In various studies,
Thermotoga maritima (Encap) encapsulins were used as a delivery system for both fluorescent probe and therapeutic drug when the nanocompartments were functionalized to bind to SP94, a hepatocellular carcinoma (HepG2) marker
[14][44][45][14,55,56]. The Encap shell protein contains two cysteine residues (C123 and C197), while C123 is located on the outer surface of Encap, allowing for SP94 peptide conjugation to the encapsulin surface. Then, a fluorescent probe fluorescein-5-maleimide was added to the shell surface. Thus, the obtained nanodevice was able to specifically bind to cells and visualize SP94 simultaneously due to the presence of a fluorophore. It was further demonstrated that a prodrug aldoxorubicin ((6-maleimidocaproyl) doxorubicin hydrazone) could be loaded into the designed construct and released under acidic conditions inside tumor cells. The dose-dependent cytotoxicity of the drug against HepG2 cells was also demonstrated.
Encapsulins can be successfully modified with photo-switchable fluorophores
[46][57]. The
Brevibacterium linens encapsulin was functionalized with spiropyran-based fluorophores (organic compounds with photochromic properties) via the carbodiimide method
[47][58]. Upon irradiation with ultraviolet and visible light, spiropyrans are able to switch between their fluorescent merocyanine photo-isomer and nonradiative isomer, allowing them to “turn on” and “turn off” fluorescence
[47][58]. The authors demonstrated that modified encapsulins carrying spiropyran molecules on their shell remained stable through at least five cycles of photo-switching.
Modification of encapsulins with fluorescent dyes allows for investigating their intracellular fate and in vivo distribution, enabling a further understanding of their biological role and function. The possibility to switch the fluorescence on and off can significantly increase imaging resolution when using such super-resolution microscopy techniques as stochastic optical reconstruction microscopy (STORM) and photoactivated localization microscopy (PALM).
In particular, spiropyran derivatives are widely used in live-cell imaging
[48][59]. The sufficient brightness and versatile chemical modification of spiropyran-based compounds allow the attachment of multiple fluorophore molecules to the encapsulin shell
[49][60]. The high sensitivity of spiropyrans to redox changes as well as to changes in temperature and pH make these molecular switchers an emerging tool in the development of fluorescent biosensors
[50][61]. Moreover, due to their biocompatibility and ability to penetrate cells, encapsulin-based fluorescent probes may also facilitate cell labeling and tracking. Simultaneous encapsulin modification with fluorescent labels and targeting moieties might also lead to the creation of an encapsulin-based targeted imaging probe. In another study
[51][36],
Brevibacterium linens encapsulins loaded with teal fluorescent protein (TFP) were found to be successfully captured by J774 murine macrophages in cell culture in vitro, yielding the intense fluorescence of macrophages. In this case, encapsulins remain within the cytoplasm without entering the nucleus. The authors note that the selected fluorescent model cargo can be replaced by a therapeutic agent and an encapsulin-based platform can be used for its delivery.
It was also shown that
Thermotoga maritima encapsulins could be genetically modified to express the FcBP peptide (IgG-Fc domain-binding peptide)
[52][62], which exhibits a high affinity to the rabbit IgG-Fc fragment
[53][63]. Indeed, real-time quartz crystal microbalance (QCM) and surface plasmon resonance (SPR) analysis indicated that FcBP is displayed on the outer surface of encapsulin and is available for binding by rabbit IgG-Fc.
While speaking about targeted delivery systems, it is worth mentioning that the encapsulin shell surface of
Rhodococcus erythropolis N771, described above, can be successfully coated with polyethylene glycol (PEG). PEG is well known as a biocompatible modifier of drug carriers, making them undetectable by the cells of the monocytic macrophage system (mononuclear phagocyte system) and reducing aggregation. The authors also showed that PEGylation does not affect the self-assembly of the nanocompartment
[21].
In one study,
Thermotoga maritima encapsulins were used as carriers of the influenza A virus protein M2 ectodomain (M2e epitope)
[54][64]. The M2 protein forms ion channels on the virion surface and is essential for the transport of viral ribonucleoprotein complexes into the cytoplasm of the host cell. The amino acid sequence of the M2 protein is highly conservative, and its immunogenicity during natural infection is quite low
[55][65]. These properties make M2e a good broad-spectrum vaccine candidate. In this work, anti-M2e epitope specific antibodies were determined in mouse serum following immunization with the obtained constructs.
Due to their size, protein compartments excellently mimic the defined intracellular environment and allow for studying the enzyme kinetics in more natural conditions
[56][57][66,67]. It was shown that non-native proteins such as GFP and firefly luciferase (Luc) could be loaded into the encapsulin shell of
Rhodococcus erythropolis [17]. Moreover, GFP retains its fluorescence properties, and luciferase showed enzymatic activity towards its substrate, luciferin. A similar study was conducted with
Brevibacterium linens encapsulins. The C-terminal sequence of the native DyP cargo protein was fused to the C-terminus of TFP
[58][68]. After confirming the structural integrity of the isolated nanocompartments, one encapsulin was found to contain 12 TFP molecules on average.
Interestingly, the encapsulins can be disassembled into protein subunits at acidic pH, loaded with a cargo of interest due to the affinity to certain protein sequences of the membrane, and finally reassembled under neutral pH conditions. Thus, molecules/particles with a size exceeding the nanocompartment pore diameter can be loaded into the encapsulin cavity. Thus, the authors of
[59][69] chose gold nanoparticles with a diameter of 13 ± 1 nm, coated with (11-mercaptoundecyl)-N,N,N-trimethylammonium bromide, as cargo. These nanoparticles were loaded into the
Thermotoga maritima encapsulin cavity by partially replacing the nanoparticle-stabilizing ligand with CLP.
This approach could be potentially applied for photothermal therapy of tumors, requiring the introduction of stable and biocompatible gold nanoparticles into the region of interest, followed by irradiation with the required wavelength and subsequent death of tumor cells, which are more sensitive to temperature increase compared to healthy cells.