Proliferating cell nuclear antigen (PCNA) is an essential factor in DNA replication and repair. It forms a homotrimeric ring that embraces the DNA and slides along it, anchoring DNA polymerases and other DNA editing enzymes. It also interacts with regulatory proteins through a sequence motif known as PCNA Interacting Protein box (PIP-box). We here review the latest contributions to knowledge regarding the structure-function relationships in human PCNA, particularly the mechanism of sliding, and of the molecular recognition of canonical and non-canonical PIP motifs. The unique binding mode of the oncogene p15 is described in detail, and the implications of the recently discovered structure of PCNA bound to polymerase δ are discussed. The study of the post-translational modifications of PCNA and its partners may yield therapeutic opportunities in cancer treatment, in addition to illuminating the way PCNA coordinates the dynamic exchange of its many partners in DNA replication and repair.
- PCNA
- structure
- protein interactions
- DNA sliding
- molecular recognition
- DNA replication
- DNA repair
1. DNA Sliding Clamps
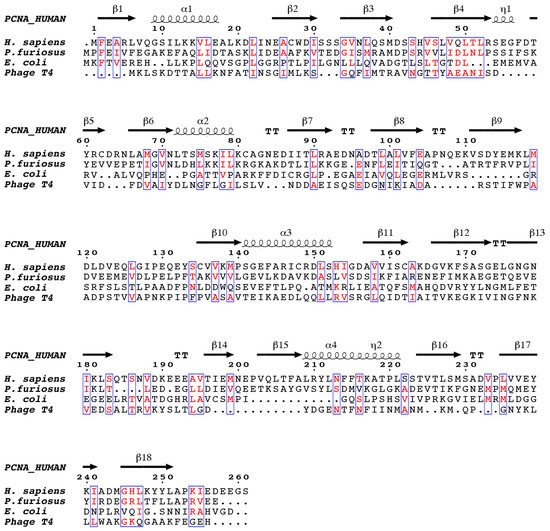
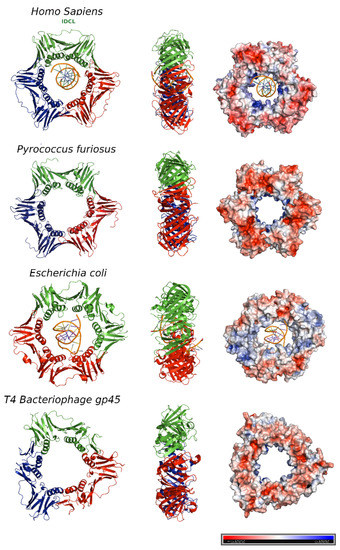
2. Human PCNA Structure
Proliferating cell nuclear antigen (PCNA) was concurrently discovered by two different groups. On one hand, Miyachi et al. [17] detected an auto-antigen in the sera of some patients with systemic lupus erythematosus, and because the protein was detected in the nuclei of dividing cells, they named it PCNA [18]. On the other hand, Bravo and Celis [19] identified a protein which was synthesized during the S-phase cell cycle and called it cyclin. Further experiments showed that both were the same protein of 29 kDa, which behaves as a homotrimer in solution. The PCNA 87 kDa ring is opened by the clamp loader Replication Factor C and is placed encircling the DNA duplex in a process where ATP is hydrolyzed [20]. Extensive studies demonstrate that PCNA is an auxiliary factor for the replication polymerases δ and ε (Pol δ and Pol ε), increasing their processivity by tethering them and sliding along the double-stranded DNA helix. Especially important is its role in the synthesis of the lagging strand, acting as a platform where Pol δ, flap endonuclease 1 (FEN1), and DNA ligase I (LIG1) bind to synthesize, process, and join Okazaki fragments [21]. PCNA recruits other factors to the replication fork, participating in DNA repair (translesion synthesis, homologous recombination, mismatch repair, and nucleotide excision repair), chromatin remodeling, and cell cycle control [21]. Not all of its partners bind simultaneously to PCNA, and switching may be triggered by different mechanisms: affinity-competition, proteolysis, or post-translational modifications [2]. The first crystal structure of human PCNA was solved in 1996, bound to the C-terminal region of the cell-cycle checkpoint protein p21WAF1/CIP1 (PDB entry: 1AXC) [22]. However, it was not until 2004 that two structures of the human PCNA trimer alone were elucidated (PDB entries 1VYM and 1W60), corresponding to two distinct spatial groups [23]. Both structures are essentially the same but a bucking trimer is seen in one, while the other presents a planar ring, indicating a certain degree of plasticity in the spatial arrangement of the PCNA trimer. There are two distinct faces of the ring: the front face, also known as the C-face (since the carboxy-terminal ends of the protomers are located there), contains a hydrophobic pocket next to the IDCL on each protomer, where polymerases and other proteins bind; while the back face has prominent loops emerging into the solvent, and is the target for post-translational modifications that alter the clamp’s properties [6]. The solution NMR (Nuclear Magnetic Resonance) spectrum of PCNA was assigned [24], and the backbone chemical shifts and many long range NOEs (Nuclear Overhauser Effect) were measured [25]. These data were consistent with the crystallographic results, indicating that the structure in solution is the same. Moreover, size exclusion chromatography coupled to multiangle light scattering (SEC-MALS) measurements showed that the trimeric form is predominant even at high concentrations, confirming that recombinant PCNA behaves as one single trimer in solution [25]. Experiments in intact cells and cell extracts showed that PCNA can form dimers of trimers [26], but even if PCNA may exist transiently as a loosely bound dimer of trimers inside cells, its functional role is unclear. Thermal and chemical denaturation studies indicate that although human PCNA has the same three-dimensional structure as the S. cerevisiae homolog, it is less stable. Human PCNA also displays increased backbone dynamics compared with yeast PCNA, especially at the helices that line the inner surface of the ring. This highly dynamic and plastic behavior of human PCNA could be an evolutionary advantage to facilitate binding to a large number of ligands [25]. The crystal structure of human PCNA bound to a 10 bp DNA duplex shows the DNA inside the channel, tilted by an angle of 15° relative to the three-fold rotation axis of the ring [27]. The PCNA-DNA interface comprises six conserved basic residues spread along five α-helices of two protomers that establish polar contacts with five consecutive phosphates of one of the DNA strands. NMR studies confirmed that primed DNA does not bind the hydrophobic pocket of PCNA.3. PCNA Binding Proteins
A large network of proteins is responsible for replicating DNA with high fidelity and for repairing DNA damage through different pathways. PCNA is a global hub in DNA metabolism that interacts with a large number of proteins involved in a variety of DNA-related processes [34][28]. As PCNA is a symmetric homotrimer in solution, it has three identical hydrophobic pockets to simultaneously bind different partners and coordinate a variety of functions in space and time [35][29]. Most of the proteins that bind PCNA either are IDPs (intrinsically disordered proteins) or have IDRs (intrinsically disordered regions). IDPs lack defined secondary and tertiary structures under physiological conditions. Although they fulfil important biological functions across all domains of life, they are more abundant in eukaryotes [36][30]. In particular, most transcription factors, as well as proteins involved in signal transduction, in eukaryotic organisms are predicted to be disordered or to contain disordered regions [37][31]. This reveals a correlation between complex cell regulation and the greater presence of IDPs/IDRs [38][32]. Moreover, most of the proteins associated with cancer have been identified as IDP or IDR-containing proteins (79%), which underlines the crucial roles they play in several cellular events that are altered in cancer, such as cell proliferation, DNA repair, and apoptosis [39][33]. The evolutionary advantage of IDPs likely lies in their plasticity that allows them to interact with many ligands and their vast regulation through post-translational modifications [40][34]. Disordered proteins have large accessible surface areas, which increase their ability to interact with diverse binding partners through short linear motifs (SLiMs) [41][35]. Many of the proteins that interact with PCNA display a characteristic SLiM known as PIP box (PCNA interacting protein-box) or an extended version called PIP degron. The canonical PIP box motif is QXXhXXaa, where h is an aliphatic hydrophobic residue (frequently I, L, or M) and a is an aromatic hydrophobic one (F, W, or Y), whereas X can be any amino acid [9]. The PIP degron motif targets PCNA for degradation and also harbors a basic residue (K or R) four amino acids after the second aromatic residue, as well as a TD motif just before the aromatic residues [42][36]. Additionally, a novel motif was identified and found to be present in several PCNA interacting proteins, named KA box, that consists of residues K-A-(A/L/I)-(A/L/Q)-x-x-(L/V) [43,44][37][38]. Structural studies of PIP box containing proteins or derived peptides in complex with human PCNA have unveiled the molecular details of the PIP motif-PCNA interface. The crystal structure of a p21 fragment bound to human PCNA was the first structural characterization of this interface (PDB code: 1AXC) [22]. Since then, several co-crystal structures of PCNA with different ligands have been solved. Overall, all PCNA interacting proteins adopt a similar conformation, which consists of an extended N-terminal region, a 310 helical turn of four residues enclosed by the hydrophobic residues of the PIP box, and a C-terminal region of variable length that sometimes adopts a β strand secondary structure and interacts with the IDCL. The conserved helix inserts into the hydrophobic pocket of PCNA, whereas the glutamine sticks into the so-called Q-pocket, establishing hydrogen bonds with the backbone of PCNA [22,42][22][36]. The assignment of the human PCNA NMR spectrum [24] allowed the analysis of the 1H-15N correlation spectra of PCNA in the presence of different ligands and the calculation of the perturbations in the NMR signals. Thereby, the NMR data provided complementary or new information regarding these interactions in solution at the residue level [45][39]. The available structural information of PCNA in complex with different partners reveals that three ligands are able to bind simultaneously the three identical protomers of the PCNA ring. Isothermal titration calorimetry (ITC) data with different peptides are all well fitted with a model assuming one set of equivalent sites, with no evidence of binding cooperativity. Therefore, it seems that ligands compete for binding with PCNA based on their affinities, which can be modulated by post-translational modifications [9]. The intrinsically disordered protein p21 is one of the PCNA binding partners showing the highest affinity (Table 1), probably because it needs to displace other proteins to block replication in response to DNA damage. This high affinity relies on efficient hydrophobic packing, as well as electrostatic interactions with the C-terminal region of PCNA [46][40]. Studies with variable length peptides of p21 suggest that basic residues at the N and C-termini encompassing the PIP box protein also contribute to an increase in the binding affinity. A recent study combining experimental and computational approaches confirmed this correlation between positive patches of residues flanking the PIP box motif and a strong affinity for PCNA [41][35]. Moreover, FEN1 endonuclease and p68 (also known as p66, the third subunit of Pol δ) exhibit fewer basic residues than p21, and accordingly present lower affinity (Table 1). However, the comparison of the structure of PCNA with full length FEN1 and with a 20-residue long PIP fragment reveals additional contacts involving the core domain and the C-terminal region of FEN1, which increase the affinity of FEN1 by three orders of magnitude.| Modification | Target Residue | Enzyme | Function | Ref. |
|---|---|---|---|---|
| Monoubiquitination | K164 | Rad18, RNF8 CRL4Cdt2 |
Promotes DNA synthesis at damaged sites | [65][41] |
| K117 | Unknown | Unknown | [69][42] | |
| Polyubiquitination | K164 | Rad5 HLTF/SHPRH |
Promotes TS | [66][43] |
| Acetylation | K13,K14, K77, K80 | CPB/p300 | Promotes PCNA degradation after NER | [31][44] |
| SUMOylation | K164 | UBC9(E2) | Inhibits DSBs formation | [68][45] |
| K254 | Unknown | Unknown | [72][46] | |
| ISGylation | K164, K168 | EFP | Terminates TLS | [70][47] |
| NEDDylation | K164 * | Rad18 | Regulates Pol η recruitment in DDR pathway |
[71][48] |
| Phosphorylation | Y114 * | Unknown | Promotes adipogenesis in response to fatty diet | [76][49] |
| Y211 | EGR | Protects PCNA from degradation and inhibits MMR | [75][50] [77][51] |
|
| Methylation | K248 | SETD8 | Promotes maturation of Okazaki fragments | [72][46] |
| Di-methylation | K110 * | EZH2 | Promotes DNA synthesis by Pol δ | [73][52] |
References
- Kuriyan, J. Sliding Clamps of DNA Polymerases. J. Mol. Biol. 1993, 234, 915–925.
- Moldovan, G.L.; Pfander, B.; Jentsch, S. PCNA, the Maestro of the Replication Fork. Cell 2007, 129, 665–679.
- Stukenberg, P.T.; Studwell-Vaughan, P.S.; O’Donnell, M. Mechanism of the sliding β-clamp of DNA polymerase III holoenzyme. J. Biol. Chem. 1991, 266, 11328–11334.
- Kelch, B.A.; Makino, D.L.; Donnell, M.O.; Kuriyan, J. Clamp loader ATPases and the evolution of DNA replication machinery. BMC Biol. 2012, 10, 1–14.
- Jeruzalmi, D.; O’Donnell, M.; Kuriyan, J. Clamp loaders and sliding clamps. Curr. Opin. Struct. Biol. 2002, 12, 217–224.
- Kelch, B.A. Review The Lord of the Rings: Structure and Mechanism of the Sliding Clamp Loader. Biopolymers 2016, 105, 532–546.
- Indiani, C.; Donnell, M.O. The replication clamp-loading machine at work in the three domains of life. Mol. Cell Biol. 2006, 7, 751–761.
- Krishna, T.S.R.; Kong, X.P.; Gary, S.; Burgers, P.M.; Kuriyan, J. Crystal structure of the eukaryotic DNA polymerase processivity factor PCNA. Cell 1994, 79, 1233–1243.
- De Biasio, A.; Blanco, F.J. Proliferating cell nuclear antigen structure and interactions: Too many partners for one dancer? Adv. Protein Chem. Struct. Biol. 2013, 91, 1–36.
- Kong, X.P.; Onrust, R.; O’Donnell, M.; Kuriyan, J. Three-dimensional structure of the β subunit of E. coli DNA polymerase III holoenzyme: A sliding DNA clamp. Cell 1992, 69, 425–437.
- Hedglin, M.; Kumar, R.; Benkovic, S.J. Replication Clamps and Clamp Loaders. Cold Spring Harb Perspect Biol. 2013, 5, 1–19.
- Matsumiya, S.; Ishino, Y. Crystal structure of an archaeal DNA sliding clamp: Proliferating cell nuclear antigen from Pyrococcus furiosus. Protein Sci. 2001, 17–23.
- Wing, R.; Drew, H.; Takano, T.; Broka, C.; Tanaka, S.; Itakura, K.; Dickerson, R.E. Crystal structure analysis of a complete turn of B-DNA. Nature 1980, 287, 755–758.
- Kelman, Z.; Donnell, M.O. Structural and functional similarities of prokaryotic and eukaryotic DNA polymerase sliding clamps. Nucleic Acids Res. 1995, 23, 3613–3620.
- Georgescu, R.E.; Kim, S.S.; Yurieva, O.; Kuriyan, J.; Kong, X.P.; O’Donnell, M. Structure of a Sliding Clamp on DNA. Cell 2008, 132, 43–54.
- McNally, R.; Bowman, G.D.; Goedken, E.R.; O’Donnell, M.; Kuriyan, J. Analysis of the role of PCNA-DNA contacts during clamp loading. BMC Struct. Biol. 2010, 10.
- Miyachi, K.; Fritzler, M.J.; Tan, E.M. Autoantibody to a Nuclear Antigen in Proliferating Cells. J. Immunol. 1978, 121, 2228.
- Kelman, Z. PCNA: Structure, functions and interactions. Oncogene 1997, 14, 629–640.
- Bravo, R.; Celis, J.E. A search for differential polypeptide synthesis throughout the cell cycle of hela cells. J. Cell Biol. 1980, 84, 795–802.
- Tsurimoto, T.; Stillman, B. Replication factors required for SV40 DNA replication in vitro. I. DNA structure-specific recognition of a primer-template junction by eukaryotic DNA polymerases and their accessory proteins. J. Biol. Chem. 1991, 266, 1950–1960.
- Slade, D. Maneuvers on PCNA Rings during DNA Replication and Repair. Genes 2018, 9, 416.
- Gulbis, J.M.; Kelman, Z.; Hurwitz, J.; O’Donnell, M.; Kuriyan, J. Structure of the C-terminal region of p21(WAF1/CIP1) complexed with human PCNA. Cell 1996, 87, 297–306.
- Kontopidis, G.; Wu, S.; Zheleva, D.I.; Taylor, P.; Mcinnes, C.; Lane, D.P.; Fischer, P.M.; Walkinshaw, M.D. Structural and biochemical studies of human proliferating cell nuclear antigen complexes provide a rationale for cyclin association and inhibitor design. Proc. Natl. Acad. Sci. 2004, 102, 1871–1876.
- Sánchez, R.; Torres, D.; Prieto, J.; Blanco, F.J.; Campos-Olivas, R. Backbone assignment of human proliferating cell nuclear antigen. Biomol. NMR Assign. 2007, 1, 245–247.
- de Biasio, A.; Sánchez, R.; Prieto, J.; Villate, M.; Campos-Olivas, R.; Blanco, F.J. Reduced stability and increased dynamics in the human Proliferating Cell Nuclear Antigen (PCNA) relative to the yeast homolog. PLoS ONE 2011, 6, e16600.
- Naryzhny, S.N.; Zhao, H.; Lee, H. Proliferating cell nuclear antigen (PCNA) may function as a double homotrimer complex in the mammalian cell. J. Biol. Chem. 2005, 280, 13888–13894.
- De March, M.; Merino, N.; Barrera-Vilarmau, S.; Crehuet, R.; Onesti, S.; Blanco, F.J.; De Biasio, A. Structural basis of human PCNA sliding on DNA. Nat. Commun. 2017, 8, 13935.
- Warbrick, E. The puzzle of PCNA’s many partners. Bioessays 2000, 22, 997–1006.
- Maga, G.; Hübscher, U. Proliferating cell nuclear antigen (PCNA): A dancer with many partners. J. Cell Sci. 2003, 116, 3051–3060.
- Click, T.H.; Ganguly, D.; Chen, J. Intrinsically disordered proteins in a physics-based world. Int. J. Mol. Sci. 2010, 11, 5292–5309.
- Wright, P.E.; Dyson, H.J. Intrinsically disordered proteins in cellular signalling and regulation. Nat. Rev. Mol. Cell Biol. 2015, 16, 18–29.
- Cordeiro, T.N.; Chen, P.; De Biasio, A.; Sibille, N. Disentangling polydispersity in the PCNA−p15 PAF complex, a disordered, transient and multivalent macromolecular assembly. Nucleic Acids Res. 2017, 45, 1501–1515.
- Iakoucheva, L.M.; Brown, C.J.; Lawson, J.D.; Obradović, Z.; Dunker, A.K. Intrinsic disorder in cell-signaling and cancer-associated proteins. J. Mol. Biol. 2002, 323, 573–584.
- Dyson, H.J.; Wright, P.E. Unfolded Proteins and Protein Folding Studied by NMR. Chem. Rev. 2004, 104, 3607–3622.
- Prestel, A.; Wichmann, N.; Martins, J.M.; Marabini, R.; Kassem, N.; Broendum, S.S.; Otterlei, M.; Nielsen, O.; Willemoës, M.; Ploug, M.; et al. The PCNA interaction motifs revisited: Thinking outside the PIP-box. Cell. Mol. Life Sci. 2019, 76, 4923–4943.
- Havens, C.G.; Walter, J.C. Docking of a specialized PIP box onto chromatin-bound PCNA creates a degron for the ubiquitin ligase CRL4Cdt2. Mol. Cell 2009, 23, 1–7.
- Liang, Z.; Diamond, M.; Smith, J.A.; Schnell, M.; Daniel, R. Proliferating cell nuclear antigen is required for loading of the SMCX/KMD5C histone demethylase onto chromatin. Epigenetics Chromatin 2011, 4, 18.
- Xu, H.; Zhang, P.; Liu, L.; Lee, M.Y.W.T. A novel PCNA-binding motif identified by the panning of a random peptide display library. Biochemistry 2001, 40, 4512–4520.
- De Biasio, A.; Campos-Olivas, R.; Sánchez, R.; López-Alonso, J.P.; Pantoja-Uceda, D.; Merino, N.; Villate, M.; Martin-Garcia, J.M.; Castillo, F.; Luque, I.; et al. Proliferating cell nuclear antigen (PCNA) interactions in solution studied by NMR. PLoS ONE 2012, 7.
- Bruning, J.B.; Shamoo, Y. Structural and Thermodynamic Analysis of Human PCNA with Peptides Derived from DNA Polymerase-δ p66 Subunit and Flap Endonuclease-1. Structure 2004, 12, 2209–2219.
- Hedglin, M.; Pandey, B.; Benkovic, S.J. Characterization of human translesion DNA synthesis across a UV-induced DNA lesion. Elife 2016, 5, 1–18.
- Elia, A.E.H.; Boardman, A.P.; Wang, D.C.; Huttlin, E.L.; Everley, R.A.; Dephoure, N.; Zhou, C.; Koren, I.; Gygi, S.P.; Elledge, S.J. Quantitative Proteomic Atlas of Ubiquitination and Acetylation in the DNA Damage Response. Mol. Cell. 2015, 59, 867–881.
- Vujanovic, M.; Krietsch, J.; Raso, M.C.; Terraneo, N.; Zellweger, R.; Schmid, J.A.; Taglialatela, A.; Huang, J.W.; Holland, C.L.; Zwicky, K.; et al. Replication Fork Slowing and Reversal upon DNA Damage Require PCNA Polyubiquitination and ZRANB3 DNA Translocase Activity. Mol. Cell 2017, 67, 882–890.e5.
- Cazzalini, O.; Sommatis, S.; Tillhon, M.; Dutto, I.; Bachi, A.; Rapp, A.; Nardo, T.; Scovassi, A.I.; Necchi, D.; Cardoso, M.C.; et al. CBP and p300 acetylate PCNA to link its degradation with nucleotide excision repair synthesis. Nucleic Acids Res. 2014, 42, 8433–8448.
- Gali, H.; Juhasz, S.; Morocz, M.; Hajdu, I.; Fatyol, K.; Szukacsov, V.; Burkovics, P.; Haracska, L. Role of SUMO modification of human PCNA at stalled replication fork. Nucleic Acids Res. 2012, 40, 6049–6059.
- Takawa, M.; Cho, H.S.; Hayami, S.; Toyokawa, G.; Kogure, M.; Yamane, Y.; Iwai, Y.; Maejima, K.; Ueda, K.; Masuda, A.; et al. Histone lysine methyltransferase setd8 promotes carcinogenesis by deregulating PCNA expression. Cancer Res. 2012, 72, 3217–3227.
- Park, J.M.; Yang, S.W.; Yu, K.R.; Ka, S.H.; Lee, S.W.; Seol, J.H.; Jeon, Y.J.; Chung, C.H. Modification of PCNA by ISG15 Plays a Crucial Role in Termination of Error-Prone Translesion DNA Synthesis. Mol. Cell 2014, 54, 626–638.
- Guan, J.; Yu, S.; Zheng, X. NEDDylation antagonizes ubiquitination of proliferating cell nuclear antigen and regulates the recruitment of polymerase η in response to oxidative DNA damage. Protein Cell 2018, 9, 365–379.
- Peng, B.; Ortega, J.; Gu, L.; Chang, Z.; Li, G.M. Phosphorylation of proliferating cell nuclear antigen promotes cancer progression by activating the ATM/Akt/GSK3β/Snail signaling pathway. J. Biol. Chem. 2019, 294, 7037–7045.
- Wang, S.-C.; Nakajima, Y.; Yu, Y.-L.; Xia, W.; Chen, C.-T.; Yang, C.-C.; McIntush, E.W.; Li, L.-Y.; Hawke, D.H.; Kobayashi, R.; et al. Tyrosine phosphorylation controls PCNA function through protein stability. Nat. Cell Biol. 2006, 8, 1359–1368.
- Ortega, J.; Li, J.Y.; Lee, S.; Tong, D.; Gu, L.; Li, G.M. Phosphorylation of PCNA by EGFR inhibits mismatch repair and promotes misincorporation during DNA synthesis. Proc. Natl. Acad. Sci. USA 2015, 112, 5667–5672.
- Peng, A.; Xu, X.; Wang, C.; Yang, J.; Wang, S.; Dai, J.; Ye, L. EZH2 promotes DNA replication by stabilizing interaction of POLδ and PCNA via methylation-mediated PCNA trimerization. Epigenetics Chromatin 2018, 11.