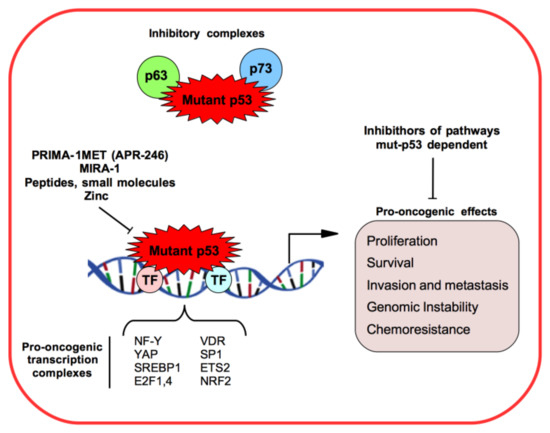
3. Mutant p53 and Long Non-Coding RNAs
Long non-coding RNAs (lncRNAs) are non-coding transcripts greater than 200 base pairs in length transcribed by RNA Pol II from independent promoters. Similar to protein-coding genes, next generation sequencing by ChIP experiments highlighted that their genomic loci are characterized by enriching H3K4 trimethylation at the transcriptional start site and H3K36 trimethylation throughout the entire gene
[69]. The lncRNA transcripts are spliced through conserved mechanisms into a mature transcript, they have fewer exons and are expressed at lower levels overall compared to protein-coding transcripts
[69]. On the basis of the ENCODE project, it is estimated that the human genome encodes more than 30,000 distinct lncRNAs, many of which are still being found and are yet to be annotated
[70]. Functionally, lncRNAs are classified as signaling, decoyed, guided, and scaffolded lncRNAs
[71]. Signaling lncRNAs are associated with specific signaling pathways, decoyed lncRNAs interact with and titrate away proteins or other RNAs, guided lncRNAs bind to the regulatory or enzymatically-active protein complexes and direct them to specific target gene promoters regulating downstream pathways and gene expressions, scaffolded lncRNAs act as a platform to which different kind of protein complexes
[71]. The lncRNAs play key roles in regulating chromatin structure, gene expression, cell growth, differentiation, and development and their mutations or dysregulation of their expression are associated with a wide range of diseases, especially tumors and neurodegenerative diseases
[72][73][72,73].
LincRNA-p21, together with other lncRNAs, was the first lncRNAs identified to be transcriptionally induced by wt-p53 ()
[74]. It served as a transcriptional repressor in the p53 pathway. The authors showed that lincRNA-p21 was able to trigger growth arrest and apoptosis through binding with hnRNP-K
[74].
Table 3. Long non coding RNAs regulated by gain of function mut-p53.
| lncRNA Name |
Cancer |
References |
| lincRNA-p21 (wt-p53) |
lung |
[74] |
| MALAT1 |
breast |
[75] | [82] |
| lincRNA-1611 |
pancreas |
[76] | [88] |
| MIR205HG |
head and neck |
[77] | [92] |
After this work, several other lines of research have correlated lncRNAs with the expression of wt-p53 in different human cancers whereas up to today few papers have documented the relationship between mutant p53 and lncRNA expression ()
[78][79][75,76].
Of considerable interest, MALAT1, also known as nuclear-enriched abundant transcript 2 or alpha, is a broadly expressed lncRNA with a length of ~8000 nt. Many studies reported that MALAT1 is a nuclear lncRNA variably expressed in many types of cancers such as endometrial, breast, cervical, and lung cancers
[80][81][82][83][77,78,79,80]. The overexpression of MALAT1 is related to hyperproliferation and metastasis
[84][81]. Recent studies show that MALAT1 regulates alternative RNA splicing of endogenous target genes using SR protein phosphorylation levels or by interacting with splicing factors such as serine-/arginine-rich splicing factor 1 (SRSF1) and SRSF3
[75][82]. It was demonstrated in basal-like breast cancers that oncogenic splicing factor SRSF1 bridges MALAT1 to mutant p53 and ID4 transcription factors
[75][82]. mut-p53 and ID4 take away MALAT1 from nuclear speckles and favor its recruitment onto the chromatin. This enables aberrant binding of MALAT1 on VEGFA pre-mRNA and modulation of VEGFA isoforms expression
[75][82].
Furthermore, MALAT1 overexpression has been reported to induce the deacetylation activity of SIRT1 and to decrease the p53 acetylation level. In this way, lncRNA suppresses the transcription levels of p53 target genes (p21, Bax, Puma, Stat3, Cyclin D, and Cyclin E), increasing the proliferation rate of the cells
[85][83]. However, considering that many post-transcriptional regulatory mechanisms of wt-p53 have been found to also be common in mut-p53, we cannot exclude that even in MALAT1 overexpressed tumors, the mechanism of regulation of mut-p53 homeostasis expression can be conserved.
Human pancreatic ductal adenocarcinoma (PDAC) is one of the most aggressive solid malignancies characterized by insensitivity to current therapy, metastasis, and poor prognosis, with a 5-year overall survival rate of less than 5%
[86][84]. It has been shown that mut-p53 proteins promote tumor proliferation, invasion, metastasis, and chemoresistance in PDAC
[87][88][89][85,86,87]. Interestingly, lincRNA-1611 was found to have significantly high expression in 22 out of 26 freshly resected human PDAC tissues, compared to normal pancreatic tissues, also displaying a positive correlation with mutations in the
TP53 gene ()
[76][88]. These data suggested that lincRNA-1611 may be used as new biomarker of PDAC tumor progression and in the future as a potential target in the personalized medicine.
Head and neck squamous cell carcinomas (HNSCCs) arise in the mucosal linings of the upper aerodigestive tract, are heterogeneous in nature and present the highest levels of tumor immune infiltration among solid cancers
[90][89]. DNA sequencing data have revealed that mutation in the
TP53 gene is frequent in HNSCC, occurring in up to 85% of human papillomavirus (HPV)-negative primary tumors, and where
TP53 mutations are associated with poor response to the radio- and/or chemo- therapies and decreased survival
[51][91][92][50,90,91]. It was reported that mut-p53, forming oncogenic complexes with NF-Y and E2F1, binds to the
MIR205HG gene promoter and positively regulates its transcription (). By splicing of MIR205HG pre-mRNA, two types of functionally-independent RNAs are produced—MIR205HG lncRNA and miR-205-5p
[77][92].
In HNSCC cancer cells, MIR205HG binds and sequesters endogenous miR-590-3p leading to increased cyclin B, cdk1, and YAP protein expression and to the deregulated cell growth
[77][92]. In HNSCC patients, high levels of lncMIR205HG are associated with tumoral samples that depend upon the expression of mut-p53 proteins
[77][92], whereas although mut-p53 forms transcriptional oncogenic complexes with the YAP cofactor
[18], it is evident that a type of feedback is established within a mut-p53/lncRNAs/miRNAs network for the homeostasis of pro-proliferative transcriptional complexes driven by mutant p53 in cancer cells.
Although other studies describe the existence of lncRNAs in relation to the expression of mutant forms of p53, their cellular function in cancer is still unknown.