Cutibacterium acnes is a member of the skin microbiota found predominantly in regions rich in sebaceous glands. It is involved in maintaining healthy skin and has long been considered a commensal bacterium. Its involvement in various infections has led to its emergence as an opportunist pathogen. Interactions between C. acnes and the human host, including the human skin microbiota, promote the selection of C. acnes strains capable of producing several virulence factors that increase inflammatory capability. This pathogenic property may be related to many infectious mechanisms, such as an ability to form biofilms and the expression of putative virulence factors capable of triggering host immune responses or enabling C. acnes to adapt to its environment. During the past decade, many studies have identified and characterized several putative virulence factors potentially involved in the pathogenicity of this bacterium. These virulence factors are involved in bacterial attachment to target cells, polysaccharide-based biofilm synthesis, molecular structures mediating inflammation, and the enzymatic degradation of host tissues. C. acnes, like other skin-associated bacteria, can colonize various ecological niches other than skin. It produces several proteins or glycoproteins that could be considered to be active virulence factors, enabling the bacterium to adapt to the lipophilic environment of the pilosebaceous unit of the skin, but also to the various organs it colonizes.
- C. acnes
- inflammation
- innate immunity
- virulence factors
- characterization
1. C. acnes Characteristics
- (1)
-
The genus Propionibacterium, comprising the species P. freundenreichii, P. cyclohexanicum, P. acidifaciens, and P. australianse.
- (2)
-
The new genus Acidipropionibacterium, comprising the species A. jensenii, A. thoenii, A. acidipropionici, A. microaerophilum, A. damnosum, and A. olivae.
- (3)
-
The new genus Pseudopropionibacterium, containing a single species: P. propionicum.
- (4)
-
The new genus Cutibacterium, comprising cutaneous Propionibacterium bacteria belonging to the species C. acnes, C. avidum, C. granulosum, and C. humerusii. P. acnes has thus been renamed C. acnes [3]. Moreover, the genus C. acnes has been further subdivided into subspecies, such as C. acnes subsp. defendens [4][5][4,5], and C. acnes subsp. elongatum [6] (Figure 1).
-
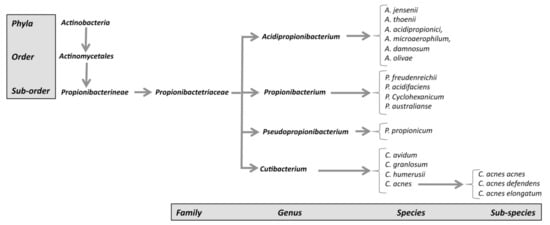 Figure 1. New classification of C. acnes strains.
Figure 1. New classification of C. acnes strains.
-
2. C. acnes Classification
| Phylotypes | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| C. acnes subspecies | WGS/Ribotyping a | SLST b | MLST8 c | CC | MLST9 d | CC | MALDI-TOF e | MLVA13 f | |
| C. acnes acnes | IA-1 | RT1, RT5 | A1-A34 | IA1 | CC1 | I-1a | CC18 | IA | IAI |
| RT532 | B1 | ||||||||
| IA-2 | RT1, RT4 | C1-C5 | IA1 | CC3 | I-1a | CC3 | IA | IA2 | |
| RT5 | |||||||||
| IB-1 | RT8 | D1-D5 | IA1 | CC4 | I-1a | CC28 | IB | IB | |
| E1-E9 | CC31 | ||||||||
| IB-2 | RT3, RT16 | F1-F14 | IA2 | CC2 | I-1b | CC28 | IB | ||
| IB-3 | RT1 | H1-H8 | IB | CC5 | I-2 | CC36 | IB | ||
| IC | RT5 | G1 | IC | CC107 | / | / | IB(IC) | / | |
| C. acnes defendens | RT2, RT6 | K1-K25 | CC6 | CC53 | |||||
| II | RT6 | II | CC30 | II | CC60 | II | II | ||
| CC71 | |||||||||
| CC72 | |||||||||
| C. acnes elongatum | III | RT9 | L1-L10 | III | CC77 | III | CC43 | III | III |