Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Nicola Tirelli and Version 2 by Rita Xu.
Yeasts are uni/multicellular eukaryotic organisms, originally thought to be ascomycetous fungi, but later recognized to also comprise basidiomycetous organisms; more typically, yeasts reproduce asexually (rapid duplication) but can also adopt sexual reproduction.
- drug delivery
- food technology
- diffusion phenomena
1. Introduction
Short recap about yeast. Yeasts are uni/multicellular eukaryotic organisms, originally thought to be ascomycetous fungi, but later recognized to also comprise basidiomycetous organisms; more typically, yeasts reproduce asexually (rapid duplication) but can also adopt sexual reproduction. A consensus definition has been proposed, and identifies yeasts “as those fungi whose asexual growth predominantly results from budding or fission, and which do not form their sexual states within or upon a fruiting body” [1]. Yeasts are best known as fermentative fungi, but it is worth noting that they are also very adaptive organisms, and have evolved the capacity to thrive (and ferment) both under aerobic and anaerobic conditions [2]. As a further testament of their capacity of adaptation, they have shown the capacity to acquire resistance to bisulfite [3] or synthetic antifungals [4][5][4,5] to the point that some yeasts can even survive nearly saturated brine solutions (e.g., Debaryomyces hansenii, also known as Candida farmata [6]). The industrial importance of these microorganisms resides in their rich metabolic activity (the very word ‘enzyme’ derives from the Greek word for leaven, ζύμη (zýmē)), which since the dawn of time has been heavily employed by humans, predominantly in food processing. Among the most popular yeasts in this business area, one could mention Saccharomyces cerevisiae and Schizosaccharomyces pombe (baking, beer making [7]), Saccharomyces bayanus (previously Saccharomyces uvarum, wine production [8]), Candida kefyr (formerly Candida pseudotropicalis, of which Kluyveromyces fragilis is the sexual stage used in dairy industry, flavors and enzymes synthesis, [9] conversion of lactose to ethanol for biofuel production [10]), and Cyberlindnera jadinii (commonly known as Candida utilis, for food flavoring [11]).
It is worth mentioning, however, that not all yeasts are beneficial; aside from the possibility for humans to develop allergic reactions [12], some yeasts are directly pathogenic such as Candida albicans (previously known as Monilia albicans; candidiasis in the gastrointestinal, reproductive and respiratory systems [13]) or Cryptococcus neoformans (a major source of lymphocytic meningitis [14][15][14,15]).
Of specific interest for this review are yeast strains capable of intracellular accumulation of large amounts of lipids, which are known as oleaginous yeasts. They can accommodate both endogenous and exogenous lipids routinely up to 20% (in some cases reportedly much more) of their weight. Examples in the literature go back decades; for example, in the 50s Cryptococcus lipofer (also known as Torulopsis lipofera) was one of the first yeasts to be shown to accumulate large amounts of sterols, [16] while in the 70s, Cutaneotrichosporon curvatum (formerly known as Candida curvata) and Cutaneotrichosporon cutaneum (also known as Basidiotrichosporon cutaneum) were first used to biotechnologically produce food-grade oil from cheese whey [17]. We refer the reader to specialized reviews for both the biochemical mechanisms presiding over lipid accumulation [18] and for the specific applications of these oleaginous yeasts [19], which range from the production of bio-diesel to that of food oils (e.g., cocoa butter substitutes). In terms of the identity of these oleaginous yeasts, possibly the most commonly employed are Cutaneotrichosporon curvatus (until recently known as Cryptococcus curvatus) [20] and Yarrowia lipolytica (formerly known as Saccharomycopsis lipolytica) [21]; a list of yeast strains with large lipid content, and therefore potentially apt for these applications, can be found in a review by Boundy-Mills [22], but it is worth noting that metabolic engineering of selected yeast types (above all of Y. lipolytic) is increasingly employed [23], as opposed to the use of a wider variety of microorganisms.
Yeast cells as microcapsules—technological advantages. Encapsulation allows for compounds of interest to be protected from a potentially aggressive environment, and released in an active form; in principle, the release occurs at specific locations and with a desired time law. A microcapsule does so at 1–1000 μm scale.
Dimensionally, yeast cells suit this definition well. For example, S. cerevisiae (baker’s yeast) cells most commonly range around 5–10 μm, always with a rather narrow size distribution; the upper end of dimensions in the yeast world is likely the Blastomyces dermatitidis, which can be as large as 40 μm [24]. Yeasts fit the definition also from a functional point of view; for a long time (1976 patent) [25], yeast cells have been known for their capacity to absorb large amounts of hydrophobes, and the process can be tuned to produce hydrophobe-loaded cells, i.e. yeast-based microcapsules (YBMCs). YBMCs have been applied to encapsulate poorly soluble actives such as flavoring agents [26], antioxidants [27], biocides (acaricides) [28], increasing their water dispersibility [29], but also providing mechanical protection thanks to the robustness of the yeast cell walls [30]. Actually, cell walls do not fulfill only a mechanical role; they are also the main barrier to both loading [31] and release. It is noteworthy that the latter (the release of actives) typically requires the presence of water, which means that in a dry state (during storage), encapsulated actives are retained with a higher efficiency [32], and release is better obtained upon wet heating [33]. Whether due to solely to these barrier properties, or maybe also to the inherent antioxidant properties of their constituents [34][35][34,35], encapsulation in yeast increases the thermo/oxidative stability of actives [36][37][36,37], considerably more than e.g. then using ‘standard’ encapsulating agents such as oligo/polysaccharides, e.g. maltodextrins [33], β-glucans [38] or modified starch, or cyclodextrins [39].
Some YBMC features can be perceived as societal advantages. Firstly, they have a sustainable and low-cost origin. Since viability of yeast is not crucial for encapsulation [31], under-utilized by-products such as spent yeast from beer making [40] can be employed. Secondly, they can be seen as safe and consumer-friendly products. Aside from the presence of yeast in virtually all bakery products, whole yeast cells, their lysates or selected components are commonly sold as super-food or flavoring agents.
The relevant literature on YBMCs is, however, poorly coherent to the point that a common parameter such as the encapsulation yield (EY%: the weight ratio between loaded compound and yeast mass) is randomly reported in relation to dry [41] or variably hydrated yeast [31][42][31,42]. A more fundamental source of variability is due to the yeast species, since different strains of the same organism might behave differently, and particular culture conditions/various levels of biological stress are likely to also have an effect.
YBMCs—commercial and medical interest. After the first seminal patent in 1976, the encapsulation of lipophilic materials in yeast cells has been protected by patents in applications related to carbonless paper, [43], fabric softeners [44], or fragrances [45] in the textile industry, nicotine for smoking cessation [46], drug delivery targeted to mucosal membranes [47], plant extracts [48], and food flavors [49]. For a more extensive review of these patents, please refer to Paramera et al. [50]. YBMCs may also have effects directly relevant to healthcare, although it is often difficult to pinpoint the precise biomolecular mechanisms and differentiate them from inflammatory (foreign-body) responses. For example, yeast cells have been linked to anti-mutagenic activity, potentially synergic with chemotherapy [51]; this may be related to the activation of inflammatory cells, which recognize yeast cells [52], phagocyte them [53] (also when in the form of yeast particles [54]), and end up exerting protective effects as a result [55]. Yeasts have also been reported to be able to trigger the opening of tight junctions in epithelial models [56], suggesting their use to control permeation through gut barriers, but this effect may also be caused by yeast triggering an inflammatory reaction: for example, C. albicans is well known to damage epithelia upon entry [57][58][57,58]. The kind of inflammatory responses elicited by YBMCs is still unclear; for example, when loaded with antigens for vaccines [59][60][61][59,60,61], siRNA [62], or even nanoparticles [63], YBMCs have produced impressive immunosuppressive effects in vivo [64], even without the use of immunosuppressive drugs [65], reportedly through combined pro-apoptotic and immunomodulatory effects [66].
2. Yeast Cells and Their Barrier Structures
Yeast cells feature an external cell wall and an inner membrane surrounding its intracellular environment, as shown in Figure 1.
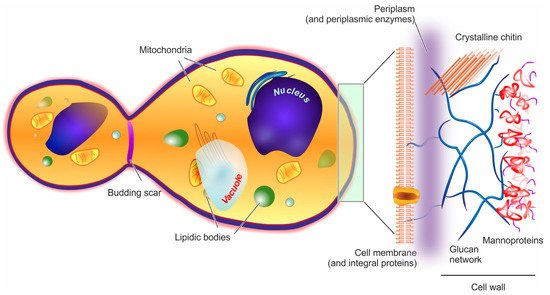
Figure 1. In the structure of a yeast cell (here represented in a post-budding state), the elements potentially acting as barriers to encapsulation (cell wall and cell membrane) are depicted in a magnified fashion on the right hand side of the figure.
Cell wall. In yeast, the wall accounts for approx. 15–20% of the cell dry mass. Its thickness is very variable (70–200 nm), since it increases in response to compression or osmotic forces [67]; structurally, however, it is always a highly polar double-layered matrix (a kind of hydrogel). Its inner part is mainly composed of branched β-(1,3) and β-(1,6) glucans (about 50% of the overall wall) [68] hydrogen-bonded to 3–4% of mostly crystalline chitin [69]. This inner layer is likely to be the main contributor to the overall mechanical resistance of the whole cell wall [70]. The outer layer consists chiefly of mannoproteins, which are negatively charged proteins highly N- or O-glycosylated with mannose or mannosyl phosphate residues linked by 1,2-, 1,3-, 1,4- and 1,6-α-linkages (the latter mostly composed of short chains typically referred to as mannan) [71]. Mannoproteins are the main contributors to the cell wall surface properties, for example, the anionic mannosyl phosphate residues determine the yeast cell surface charge [72] and their reduction in number determines an increase in its hydrophobicity [73]. Furthermore, through their covalent linkage to the β-glucan layer, mannoproteins contribute to the wall outer porosity [74], and above all to yeast adhesion, for example, the C. albicans adhesins that allow its binding to oral epithelial cells are all mannoproteins [75].
Of note, the overall wall composition is rather constant throughout the different yeast species [76], and that of S. cerevisiae can be considered as representative for almost all Ascomycetes [77]; it must be pointed out, however, that although the main components are preserved, the actual detailed composition is not at all simple, with about 1200 genes having being linked to a role in the cell wall [78].
Plasma membrane. It is composed in equal parts of lipids (mainly glycerophospholipids and fatty acids, with a smaller quantity of sterols such as ergosterol and sphingolipids) and proteins, and is connected by glycoproteins and glycolipids to the cell wall, from which it is separated by a non-continuous, enzyme-rich region known as the periplasm. The membrane main functional role is the regulation of the transport from/to the cell. [79] In the outer part of the membrane, phosphatidylcholine, phosphatidylethanolamine, phosphatidic acid, and sphingolipids are mostly found, while the more negatively charged phosphatidylserine and phosphatidylinositol are mostly present only in the inner membrane [70]; as in mammalian cells, lipid rafts are commonly observed, and have important roles in protein segregation and localization [80], which in turn have crucial effects in cell growth and death [81], or mating and division [82].
Intracellular ultrastructure. The yeast interior is compartmented into a nucleus, one or, more commonly, several vacuoles, mitochondria, the endoplasmic reticulum, and a number of vesicular bodies that include peroxisomes and oxisomes. It is worth noting that vacuoles are large (sometimes similar in size to nuclei), mildly acidic (pH 6–6.5) vesicles, whose functions in both storage and stress response/digestion bear some functional analogies to mammalian lysosomes/autophagosomes [83][84][83,84].