Alcoholic liver disease (ALD) is one type of liver disease, causing a global healthcare problem and mortality. The liver undergoes tissue damage by chronic alcohol consumption because it is the main site for metabolism of ethanol. Chronic alcohol exposure progresses from alcoholic fatty liver (AFL) to alcoholic steatohepatitis (ASH), which further lead to fibrosis, cirrhosis, and even hepatocellular cancer. Therapeutic interventions to combat ALD are very limited such as use of corticosteroids. However, these therapeutic drugs are not effective for long-term usage. Therefore, additional effective and safe therapies to cope with ALD are urgently needed. Previous studies confirmed that edible food plants and their bioactive compounds exert a protective effect against ALD.
- alcoholic liver disease
- bioactive compounds
- polyphenols
- antioxidants
- gut microbiota
Note: The following contents are extract from your paper. The entry will be online only after author check and submit it.
1. ALD: Epidemiology and Risk Factors
1.1. Alcohol Metabolism
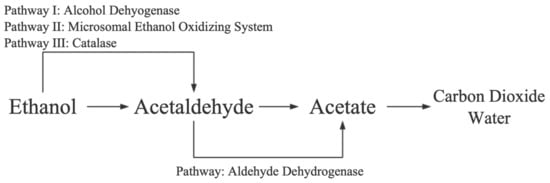
1.2. Mechanisms of Alcoholic Liver Disease
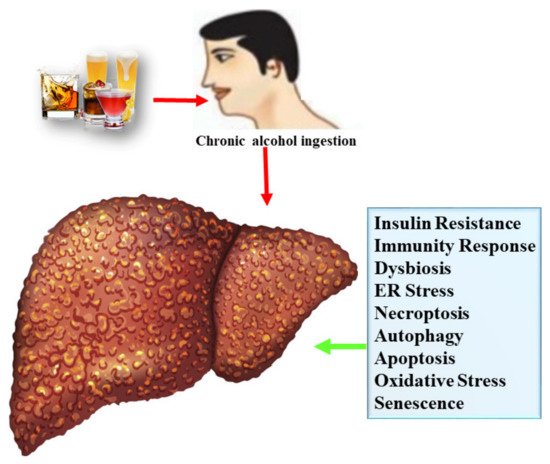
1.2.1. Lipogenesis
1.2.2. Oxidative Stress
1.2.3. Inflammatory Response
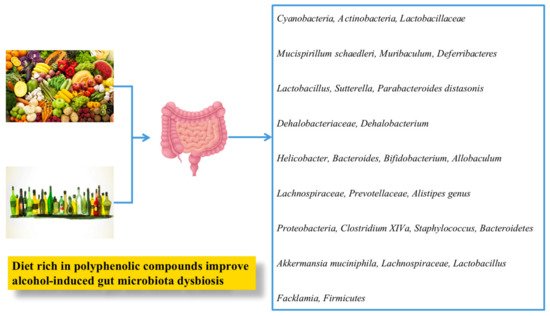
1.2.4. Gut Microflora
1.2.5. Endoplasmic Reticulum Stress
1.2.6. Apoptosis
2. Multicomponent Food Plant Extracts against ALD
2.1. Fruits
2.2. Vegetables
2.3. Spices
2.4. Cereals and Grains
2.5. Tea and Coffee
| Edible Food Plant Category | Source | Bioactive Compounds | Study Design | Major Findings | Ref. | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fruits | Blueberry | ND | Blueberry juice combined with mixed probiotics containing | (Bifidobacterium | , | Lactobacillus bulgaricus | , and | Streptococcus thermophilus | ; blueberry juice: 1.5 mL/100 g; 20 mL/100 g probiotics) for 10 days were given to ethanol-induced mice. | Blueberry juice and probiotics increased SOD, GSH, and HDL-C levels, decreased AST, ALT, TG, TC, LDL-C, and MDA, suppressed acetylated FOXO1, FOXO1, FasL, and caspase-3, and increased the SIRT1 in ethanol-exposed mice. | [69] | [110] | ||
| Mango | Mangiferin | Mangiferin (50 and 100 mg/kg bw) was orally given to ethanol-exposed rats for 12 weeks. | Mangiferin effectively regulated metal elements and FFA in serum, modulated specific alcohol-hepatitis-related genes, metabolic pathways, and potential biomarkers in alcoholic hepatitis rats. | [67] | [144] | |||||||||
| Grape | Quercetin, myricetin, rosmarinic acid, catechin, b-type procyanidin trimer, caffeic acid-O-hexoside, epicatechin | Grape-leaf extract (250–500 mg/kg) was orally given to ethanol-induced rats for 12 days. | Grape leaf extract attenuated liver injury by improving antioxidant activities, suppressed NF-κB p65 and proinflammatory cytokines (TNF-α), and normalized histopathological changes in liver. | [68] | [145] | |||||||||
| Pomegranate | ND | Pomegranate (600 mg/kg bw) was orally given to ethanol-induced female Fischer wild-type rats for 10 days. | Pomegranate pretreatment markedly reduced alcohol-mediated plasma endotoxin, gut barrier dysfunction, and inflammatory biomarkers and inhibited elevated oxidative and nitrative stress marker proteins. Moreover, pomegranate also restored the levels of intestinal tight junction proteins (claundin-3, ZO-1, occludin, and claudin-1). | [72] | [148] | |||||||||
| Cranberry | Cyanidin 3-O-galactoside, peonidin 3-O-galactoside and peonidin 3-O-arabinoside, (+)-catechin, (−)-epicatechin and (−)-epicatechin 3-gallate, procyanidin oligomers, myricetin aglycone, quercetin derivatives, benzoic acid, hydroxycinnamic acid derivatives, and hydroxybenzoic acids | Male albino Wistar rats were received cranberry polyphenols daily, 4 mg/kg bw, along with 4 g/kg bw for 8 weeks | Cranberry polyphenols ameliorated alcoholic liver damage and hepatic steatosis, decreased TG, AST, and ALT activities, diminished TNF-α, TGF-β levels, and free radical generation in mitochondria during intoxication. | [73] | [149] | |||||||||
| Wolfberry | Zeaxanthin dipalmitate | BRL-3A cells were treated with ethanol (250 mM) or Wolfberry-derived zeaxanthin dipalmitate (1 µM). Wolfberry-derived zeaxanthin dipalmitate (10 mg/kg bw) was administered to ethanol-induced rats for 4 weeks. |
Wolfberry-derived zeaxanthin dipalmitate attenuated hepatocyte and whole-liver injury in both ethanol-treated cells and rat model. The underlying mechanism was mainly due to Wolfberry-derived zeaxanthin dipalmitate directly targeted on cell membrane and including receptor P2 × 7 and adipoR1 which further modulate PI3K/AMP-FoXO3 pathways to restore mitochondrial autophagy. Moreover, WZD also alleviates hepatic inflammation by suppressing NLRP3 inflammasome. | [74] | [150] | |||||||||
| Mango | Mangiferin | Mangiferin (100 and 200 mg/kg bw) was orally given to ethanol-exposed rats for 11 days. | Mangiferin attenuated liver injury induced by chronic plus a single binge ethanol by restoring PDE3B stability, which further activated the AMPK/TBK1 signaling and inhibited NF-κB activation, leading to decreased FFA. | [75] | [151] | |||||||||
| Lychee | Procyanidin B2, quercetin, 3-O-rutinoside-7-O-a-L-rhamnosidase, isorhamnetin-3-O-rutinoside, (−)-epicatechin, rutin | Lychee pulp (0.4 to 0.8 g/L) was given to mice along with ethanol-containing liquid diet (4%) for 8 weeks. | Lychee pulp ameliorated ALD by decreasing TG, improved the antioxidant status, reduced Nrf2, suppressed lipid synthesis genes, elevated fatty acid β-oxidation expression, and decreased the serum endotoxin level. | [77] | [153] | |||||||||
| Lychee | Lychee pulp (0.2 and 0.4 g/kg bw) was given to mice along with ethanol-containing liquid diet for 8 weeks. | Lychee pulp supplementation decreased ALT and AST levels, inhibited serum and hepatic oxidative stress, suppressed mitochondrial 8-hydroxy-2’-deoxyguanosine level, and elevated the hepatic ATP level, mitochondrial membrane potential, activities of mitochondrial complexes I and IV, and mitochondrial DNA content. | [78] | [154] | ||||||||||
| Lychee | Lychee pulp (0.2 and 0.4 g/kg bw) was given to mice along with ethanol-containing liquid diet for 8 weeks. | Lychee pulp phenolic extract alleviated ethanol-induced liver injury in treated mice via reversed alteration of intestinal microbiota composition, downregulated inflammation markers, increased the expression of intestinal tight junction proteins, antimicrobial proteins, and mucus protecting proteins, repressed NF-κB p65, and suppressed CD14 and TLR4 expression. | [79] | [155] | ||||||||||
| Blueberry | ND | Blueberry polyphenols extract (100 and 200 mg/kg bw) was orally given to ethanol-exposed mice for 30 days. | Blueberry polyphenols decreased the TG lipid droplet content in liver and serum TG and TC levels and decreased lipogenic and increased lipodieretic mRNA levels. Blueberry polyphenols promoted autophagy to accelerate lipid metabolism and thus protect from ALD. | [80] | [156] | |||||||||
| Mulberry | Water extracts of mulberry (0.3 g/kg bw) were orally administered to chronic ethanol-induced rats. | Water extracts of mulberry decreased TG level and MDA contents, increased glycogen deposits, prevented the disruption of the hepatic cells and nuclei, and decreased Firmicutes to Bacteroidetes ratio. | [82] | [158] | ||||||||||
| Indian gooseberry | ND | Indian gooseberry was administered (250 mg/kg bw) to alcohol-exposed rats. | Indian gooseberry significantly reduced lipid peroxidation levels and restored antioxidant level. | [84] | [160] | |||||||||
| Ginseng berry | Ginsenoside F5, ginsenoside Rd, ginsenoside F3, and ginsenoside Re | Ginseng berry extract at the dosage of 0.5–5 mg/mouse along with ethanol was given to mice for 10 days. | Ginseng berry attenuated ALD by improving antioxidant level and reducing inflammatory mediators. | [85] | [161] | |||||||||
| Apricot | 3-caffeoylshikimic acid, 3-feruloylquinic acid, 3-hydroxy-3-methoxycarbonyl glutaric acid, 1,5-dimethyl citrate, 3,4,5-trimethoxyphenyl-β-D-glucopyranoside, prunate, methyl 3-caffeoylquinate, 3-O- caffeoylquinic acid |
AML-12 cells were treated with ethanol or chlorogenic acid. Apricot extract (100 mg/kg bw) along with alcohol (1 g/kg bw) was orally given to mice for 5 days. |
Chlorogenic acid derived from apricot extract ameliorated ALD in AML-12 cells by inhibiting alcohol-induced apoptosis, MAPK activation, and antioxidant activities. Apricot extract protected ALD by suppressing lipogenesis in liver tissue, inhibiting activation of SREBP-1, and suppressing hepatic apoptosis and inflammation via ROS-mediated p53 signaling pathway in mice with alcohol-induced liver injury. |
[86] | [162] | |||||||||
| Lemon | ND | Lemon juice (10 mL/kg bw) was orally given to alcohol-induced C57BL/6 mice for 15 days. | Lemon juice markedly inhibited alcohol-induced increase of ALT, AST, lipid peroxidation levels, and hepatic TG, improved antioxidant capacity (SOD and CAT), and improved histopathological changes in ALD mice. | [88] | [164] | |||||||||
| Citrus depressa | 5-O-demethylnobiletin, sinensetin, tangeretin, and nobiletin | Citrus depressa extract (300 mg/kg) was orally administered to ethanol-induced mice for 8 weeks. | Citrus depressa extract remarkably decreased AST, ALT, TNF-α levels, hepatic MDA, and CYP2E1 expression, and increased glutathione in ALD mice. | [89] | [165] | |||||||||
| Noni fruit | ND | Noni fruit was orally given to ethanol-exposed mice. | Noni fruit reversed the ethanol-induced changes in mice such as ALT, AST, gamma-glutamyl transferase, LDL-C, HDL-C, TG, and TC. | [90] | [166] | |||||||||
| Vegetables | Purple potato | Petunidin-3-glucoside, Petunidin-3-rutinoside-5-glucoside, Petunidin-3-caffeoyl-rutinoside-5-glucoside | Purple potato extract was administered at the dosage of 5 and 10 mg/kg bw to ethanol-exposed mice for 5 weeks. | Purple potato extract ameliorated ALD by decreasing ALT, AST, TG, and TC, reducing MDA contents and CYP2E1 protein expression, and increasing GSH and SOD levels in ethanol-exposed mice. | [94] | [170] | ||||||||
| Garlic oil | ND | Human normal cell LO2 was treated with ethanol (100 mM). Garlic oil was administered (50 to 200 mg/kg bw) to ethanol-exposed male Kunming mice. |
Garlic oil decreased n-SREBP-1c and CYP2E1 and increased PPAR-α protein levels in human normal cell L02. Garlic oil decreased n-SREBP-1c and CYP2E1 and increased PPAR-α protein levels in ethanol-induced mice. Additionally, garlic oil decreased FAS and inhibited ethanol-induced hepatic mitochondrial dysfunction. |
[99] | [175] | |||||||||
| Asparagus officinalis | ND | Asparagus extracts (400 mg/kg bw) were orally administered to male Wistar rats for 70 connective days. | Edible asparagus protected from toxicity mediated by alcohol by improving antioxidant status. | [102] | [178] | |||||||||
| Okra seed oil | Polyunsaturated fatty acids; ROS: reactive oxygen species; short-chain fatty acids; monounsaturated fatty acids | Okra seed oil (400 and 800 mg/kg bw) was given to mice for 8 weeks. | Okra seed oil attenuated alcohol-induced liver damage via inhibition of liver fat accumulation, decreased MDA content, decreased hepatic pro-inflammatory cytokines (IL-6, TNF-α, and IL-1), increased SOD and GSH levels, and attenuated lipid metabolic disorder. Furthermore, okra seed oil also modulated gut microbiota dysbiosis by enhancing the | Bacteroidetes | population and reducing the | Proteobacteria | proportion, | Staphylococcus | , and | Clostridium XlVa | . | [103] | [179] | |
| Artichoke | ND | Ethanolic extract of artichoke (0.4 to 1.6 g/kg) was given to ethanol-induced ICR mice for 10 days. | Artichoke remarkably attenuated ALD by preventing elevated levels of ALT, AST, TG, and TC, increased SOD and GSH, decreased MDA level, and suppressed inflammatory pathway (TLR4/NF-κB) in ethanol-induced ICR mice. | [104] | [180] | |||||||||
| Rhubarb | ND | Rhubarb extract (0.3%) was given to C57BL/6J mice for 17 days. | Rhubarb extracts protected alcohol-induced liver injury by modulating intestinal microflora, improving antioxidant level, and reducing inflammatory response. | [105] | [181] | |||||||||
| Bitter gourd | ND | Bitter gourd was administered (500 mg/kg bw) to C57BL/6 mice fed an alcohol-containing liquid diet for 30 days. | Bitter gourd supplementation reduced the steatotic alternation of liver histopathology, decreased AST, ALT, hepatic TG level, and MDA content, improved antioxidant defense system (SOD, GSH, GRd, GPx, and CAT), reduced pro-inflammatory cytokine levels (IL-6, TNF-α, and IL-1β), and suppressed ACC, CYP2E1, FAS, and SREBP-1 protein expression in alcohol-induced mice. | [106] | [182] | |||||||||
| Spices | Cinnamon | ND | Cinnamon bark extract (0.5 mL) was administered for 4 days prior to ethanol, and on 5th day, ethanol (6 g/kg bw) was administered. Murine RAW 264.7 macrophage-like cells were treated with cinnamon bark extract (4 µL). |
Cinnamon bark extract protected liver from alcohol via the inhibition of MyD88 expression both in vitro and in vivo. | [107] | [183] | ||||||||
| Fenugreek | ND | Fenugreek seed polyphenol extract (200 mg/kg bw) and ethanol (6 g/kg per day) were fed to rats for 30 days. | Fenugreek seed polyphenol extract inhibited lipid accumulation in ethanol-induced rats. | [108] | [184] | |||||||||
| Crocus sativus | L. | Safranal, crocin, myricetin, and quercetin | Crocus sativus | L. (saffron) petal extract was administered (167.5 and 335 mg/kg/day) to ethanol-induced rats for 30 days. | Saffron polyphenolic extract protected liver from ethanol by reducing inflammation in ethanol-administered rats. | [109] | [185] | |||||||
| Parsley oil | ND | Parsley oil (50 mg/kg bw) was given to adult male albino rats for 4 weeks. | Parsley oil attenuated alcohol-induced liver injury by oxidative stress mechanism. | [110] | [186] | |||||||||
| Syzygium aromaticum L | . | ND | Polyphenol-rich extract of clove buds (Clovinol) (100 mg/kg bw) was given to ethanol-induced rats for 30 days. | Clovinol decreased alcohol-associated oxidative stress and inflammatory changes in ethanol-induced rats. | [111] | [187] | ||||||||
| Thymus vulgaris | ND | Thymus vulgaris | leaves were orally given (500 mg/kg bw) to ethanol-induced rats for 21 days. | Co-administration ( | Thymus vulgaris | and ethanol) modulated several biomarkers such as ALP, AST, albumin, CAT, MDA, SOD, GST, and lipid profile. | [112] | [188] | ||||||
| Peppers | Capsaicin | Capsaicin was given (10 and 20 mg/kg) to ethanol-induced rats. | Capsaicin ameliorated alcohol-induced liver injury by modulating matrix metalloproteinases and suppressing free radical formation and oxidative stress. | [113] | [189] | |||||||||
| Cereals | Black rice | Cyanidin-3,5-diglucoside, cyanidin-3-glucoside, cyanidin-3-rutinoside, and peonidin-3-glucoside | Alcohol (3.7 g/kg bw) and anthocyanin-rich black rice extract (125, 250, and 500 mg/kg bw) dissolved in water was administered using an intragastric tube for 45 days. | Anthocyanin-rich black rice extract attenuated ALD by decreasing serum AST, ALT, TCH, TG, and GGT levels and improving antioxidant levels. | [116] | [105] | ||||||||
| Rice | Acacetin, caffeic acid, ferulic acid, sinapic acid, p-coumaric acid, quercitrin, vitexin, rutin, hesperidin, ethyl caffeate, and ethyl coumarate | Rice bran phenolic extract (0.25 or 0.50 g/L) was fed along with alcohol-containing liquid diet (4%) to mice for 8 weeks. | Anthocyanin-rich black rice extract supplementation ameliorated ALD by repressing inflammatory responses in liver, intestinal microbiota dysbiosis, and barrier dysfunction and inactivated the endotoxin-TLR4-NF-κB pathway. | [114] | [190] | |||||||||
| Rice | Acacetin, caffeic acid, ferulic acid, sinapic acid, p-coumaric acid, quercitrin, vitexin, rutin, hesperidin, ethyl caffeate, and ethyl coumarate | Rice bran phenolic extract (0.25 or 0.50 g/L) was fed along with alcohol-containing liquid diet (4%) to mice for 8 weeks. | Rice bran phenolic extract exerted protective effect against ALD in mice fed with an ethanol-containing diet via microRNAs-PGC-1α-TFAM signal pathway. |
[115] | [191] | |||||||||
| Tartary buckwheat | ND | Acute liver injury model group: buckwheat ethanol extracts (8.35, 16.70 and 41.75 mL/kg bw) and ethanol (4 g/kg bw) were intragastrically administered to rats for 7 consecutive days. Chronic alcoholic liver injury: buckwheat ethanol extracts (8.35, 16.70 and 41.75 mL/kg bw) and ethanol (3 g/kg/day bw; 37.5% volume fraction) intragastrically administered to SD rats for 8–9 consecutive weeks. |
Tartary buckwheat extract administration significantly decreased serum ALT, AST, and hepatic MDA and improved hepatic GSH level. | [117] | [192] | |||||||||
| Mung bean extract | Vitexin and isovitexin | Mung bean extract (containing 15 mg vitexin and 13 mg isovitexin, respectively, per kg bw) was given along with spirit (56% alcohol, 16 mL/kg bw) 2 h after the doses of mung bean extract for 14 days. | Mung bean extract decreased ALT and AST and improved antioxidant levels. | [122] | [197] | |||||||||
| Tea | Pu-erh tea | Gallocatechin, gallic acid, and caffeine | Pu-erh tea extract (1 or 4 g/L | w | / | v | added into drinking water) and ethanol solution (10% | w | / | v | ) were administered by gavage for 30 days. | Pu-erh tea extract contributed to the protective effect against ALD by improving oxidative stress, reducing lipid accumulation, reducing inflammation, and modulating microbiomic and metabolomic responses. | [129] | [204] |