1. Introduction
Innate immunity relies on the recognition of evolutionarily conserved microbe-specific molecules, termed pathogen-associated molecular patterns (PAMPs). Germline encoded pattern recognition receptors (PRRs) expressed on the cell surface, endosomes or in the cytosol detect and respond to these PAMPs. Although, the domains of these PRRs are highly conserved, extensive species-specific expansions and domain shuffling result in an advantage to an organism living in pathogen-rich environments. The PRRs expressed by mammalian cells are Toll-like receptors (TLRs), the NOD-like receptors (NLRs), AIM2-like receptors (ALRs), RIG-like receptors (RLRs) and C-type lectin receptors (CLRs), with each family member recognizing specific molecular signatures [1]. Two of these families of PRRs are conserved from early invertebrates to mammals: the transmembrane TLRs and the intracellular NLRs [2][3][2,3].
Our skin acts as a sentinel organ, determining when and how to respond to a broad range of environmental insults during both homeostatic and pathologic situations. The skin forms a physical barrier through the cornified envelope of stratum corneum and via tight-junctions in lower layers, a chemical barrier by maintaining an acidic pH and antimicrobial peptide expression and finally, there is the immunologic barrier formed by keratinocytes and infiltrating immune cells of both the innate and adaptive immune systems [4]. These layers of barriers interact with each other to protect the organism from harmful stimuli. Keratinocytes are the main cell type of the epidermis and as immunocompetent cells are implicated in the protection against harmful threats, by the expression of a wide range of PRRs, including TLRs and NLRs [5][6][7][5,6,7]. The activation of PRRs induces keratinocytes to express antimicrobial peptides and immune mediators, which promote the recruitment of professional immune cells [4]. Murine and human TLRs in skin biology have been discussed elsewhere [5][6][5,6], here we will focus on the NLR family and discuss its role in the immune defence in the skin.
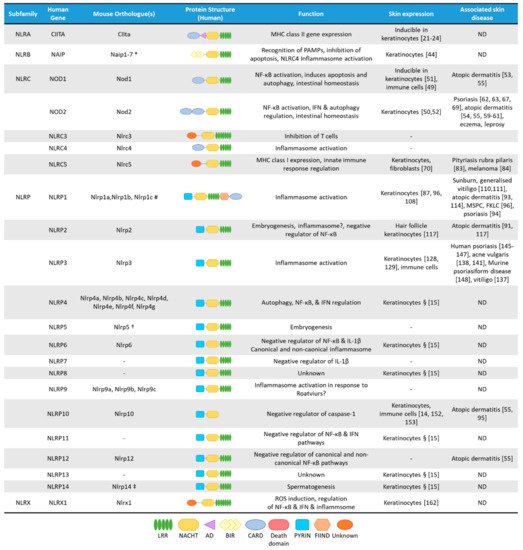
NLRs are cytosolic receptors widely identified in non-vertebrates and vertebrates, and have functional analogues, the R-proteins, in plants [8]. The number of NLR genes differ from species to species: humans express 23 NLR family members, while mice have at least 34 NLR paralogues () [1][8][1,8]. NLRs are multi-domain containing proteins, comprising of a C-terminal domain with a series of leucine rich repeats (LRRs), a central nucleotide-binding NACHT domain, and an N-terminal effector domain [9]. The N-terminal domain is variable, and NLRs are divided into five subfamilies based on their distinctive N-terminal domain: NLRAs that have an acidic activation domain, NLRBs that possess a baculovirus inhibitor of apoptosis repeat (BIR)-like domain, NLRCs that feature a caspase activation and recruitment domain (CARD) or a Death domain (DD), and the NLRP subfamily that contain a PYRIN domain [9]. The NLRX subfamily contains one member, and its nomenclature derives from an uncharacterized N-terminal domain that lacks homology with other NLR effector domains.
In the cytosol, NLRs remain in an auto-inhibitory state. The LRR domains are thought to be responsible for ligand binding but this has not been experimentally shown to date for most NLRs and this dearth of evidence has led to the belief that mammalian LRRs might not have necessarily retained this function [10][11][10,11]. The LRR region also maintains the NLR in an auto-inhibitory state, as demonstrated by the crystal structure of NLRC4, where the LRR obstructs the NACHT domain [12]. The NACHT domain possesses dNTPase activity, which governs the ATP-dependent oligomerisation. Although, the NACHT domain controls oligomerisation, additionally ligand-binding can occur in this region. Upon activation, the N-terminal domain activates distinctive downstream signalling cascades resulting in an inflammatory response. This innate immune response also serves to influence the adaptive arm of the immune system [1]. Despite subfamilies sharing the same domain, individual members can elicit different downstream effects. For example, the NLRC family that contains a CARD domain, induces inflammasome activation, regulates nuclear factor κ-light-chain-enhancer of activated B cells (NF-κB) or type I interferon (IFN) signalling pathways or engages in transcriptional regulation [13].
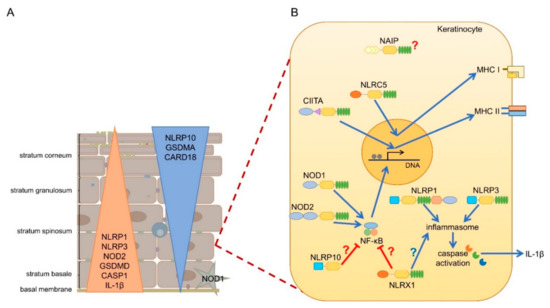
mRNA expression of most NLRs are found in the skin, but since NLR activation is a very complex process, and some NLRs are characterized by unique cell-type specific features, without their functional evaluation in keratinocytes, their functions cannot be clearly addressed. Cornification of keratinocytes also affects the expression of NLRs and their interacting partners (A) [14]. Here, we will summarize the current knowledge on epidermal NLR expression and functions and their potential contribution to skin disease.
Figure 1. The NLR superfamily. Schematic depicting the five subfamilies of human NLRs and their mouse orthologues. Additionally, depicted are protein structural domains, the known function of the NLRs, their expression in skin cells and association with skin diseases. AD, Acidic transactivating domain, BIR, baculovirus IAP repeat, CARD, caspase-activation and recruitment domain, FIIND, Function-to-find domain, FKLC, familial keratosis lichenoides chronica, IFN, Interferon, LRR, Leucine-rich repeats, MSPC, multiple self-healing palmoplantar carcinoma, NACHT, a domain found in NAIP, CIITA, HET-E and TEP1, ND, not determined. * Murine Naip3 lacks a NACHT-LRR and contains three BIR domains, Naip4 lacks a NACHT-LRR and contains one BIR domain, Naip5 & Naip6 lack an LRR region. # Murine Nlrp1a lacks a PYRIN, Nlrp1b lacks a PYRIN and FIIND, Nlrp1c lacks PYRIN, FIIND and CARD domains. † Murine Nlrp5 lacks a PYRIN domain. ‡ Murine Nlrp14 lacks a PYRIN domain. § Based on mRNA expression assessed by Reverse Transcriptase-PCR in human primary keratinocytes
[15].
Figure 2. Expression and function of NLRs in human epidermis. (A) While inflammasome forming (NLRP1, NLRP3) and pro-inflammatory (NOD1, NOD2) NLRs are expressed in basal layers of the epidermis along with other pro-inflammatory genes, anti-inflammatory NLRs (NLRP10) and inflammasome inhibiting genes (CARD18) are rather enriched in the upper layers of the epidermis. (B) CIITA and NLRC5 are inducible in keratinocytes by IFN-γ and regulates MHC II and MHC I expression, respectively. NOD1 and NOD2 is activated by bacterial products, leading to NF-κB activation and inflamma . and NLRP3 are both forming inflammasomes in keratinocytes leading to IL-1β secretion NAIP, NLRP10 and NLRX1 are expressed in keratinocytes, however their functions in keratinocytes are not described so far. NLRP10 and NLRX1 were shown to inhibit NF-κB activity in professional immune cells and regulate inflammasome activation, however, whether these functions are dominant in keratinocytes is unknown.
2. The NLRA Subfamily
The NLRA subfamily comprises of a sole member, namely: the Class II Major Histocompatibility Complex Transactivator (CIITA), which contains an
N-terminal acidic transactivation domain, but also a CARD domain, a nuclear localization signal and four LRRs
[16]. CIITA has been recognized as the “master regulator” of Major Histocompatibility Complex (MHC) class II molecule (MHC-II) expression since it controls the differential expression of MHC-II genes
[17]. CIITA also plays a role in human MHC class I (MHC-I) expression, a function that is not observed in mice
[18]. CIITA lacks a DNA-binding domain but controls transcription by recruiting the transcription machinery, including TFIID and TFIIB
[19][20][19,20]. It also induces phosphorylation of RNA polymerase I and enlists the chromatin remodelling coactivators
[19][20][19,20].
CIITA is a founding member of the NLR protein family, but for a long time, it remained detached from the rest of the protein family as the only transcriptional regulator. Although, MHC-II is typically expressed by professional antigen-presenting cells, such as dendritic cells (DCs), B cells, macrophages, and thymic epithelial cells, it is long known that expression can also be induced in keratinocytes by IFN-γ
[21][22][23][24][21,22,23,24]. Moreover, keratinocytes express MHC-II in a variety of skin disorders, including psoriasis, allergic contact dermatitis, and atopic dermatitis, which accompanies infiltration of activated T cells
[25][26][25,26].
In mice and humans, CIITA expression is regulated by three and four distinct promoters, respectively, resulting in different isotypes
[16][22][16,22]. Human keratinocytes mainly express type IV CIITA transcribed from promoter IV upon IFN-γ stimulation
[22], which can be further induced by interleukin (IL)-18
[27], which subsequently leads to MHC-II expression
[22]. MHC-II expressing human keratinocytes have been reported to present
Mycobacterium leprae antigens (hsp65) and induce the proliferation of Th
1 cells, indicating that human keratinocytes can process and present some intact protein antigens
[28], however, they fail to activate naïve T-cells
[29][30][31][29,30,31]. In contrast, mouse keratinocytes are unable to present intact proteins to specific T cells
[30]. Although keratinocytes were shown to be able to express CD86 costimulatory molecules
[32], generally they do not express adequate levels of CD80/CD86
[33][34][35][33,34,35], which may explain their inability to properly activate resting or naïve T cells.
3. The NLRB Subfamily
Similarly to NLRA, the NLRB subfamily contains a single member; the neuronal apoptosis inhibitory protein (NAIP, also called NLRB1 or BIRC1)
[9]. NAIP contains a BIR-like domain in its
N-terminus
[9]. There is a single human NAIP orthologue but mice contain at least 7 paralogues
[36]. NAIP is an anti-apoptotic protein that acts by inhibiting caspase activity by activation of mitogen-activated protein kinase (MAPK) pathways
[37][38][39][37,38,39]. NAIP exerts dual-functionality as it also forms an inflammasome with NLRC4
[40]. NAIP proteins directly interact with bacterial PAMPs, including flagellin. Human NAIP also binds to the T3SS needle protein of the bacterial type III secretion systems
[41]. NAIPs then recruit NLRC4 as an adaptor to induce caspase-1 activation, which it does by direct CARD:CARD interactions
[42][43][42,43].
According to the Human Protein Atlas, NAIP is expressed in various cell types, including keratinocytes
[44]. However, compared to other NLRs the role of NAIP in the skin is poorly studied so far. NLRC4 has not been reported to be expressed in the skin, so NAIP/NLRC4 inflammasome activation may be unlikely.
4. The NLRC Subfamily
The NLRC subfamily is characterized by the presence of an
N-terminal CARD-domain and is the second largest subfamily of NLRs, consisting of six members: nucleotide oligomerization domain 1 (NOD1/NLRC1), NOD2 (NLRC2), NLRC3, NLRC4 and NLRC5
[45], of which NOD1, NOD2 and NLRC5 are expressed in the skin and keratinocytes, to varying extents
[46].
4.1. NOD1 and NOD2
NOD1 and NOD2 are prototypic PRRs and recognize intracellular bacterial peptidoglycans motifs, having one (NOD1) or two (NOD2)
N-terminal oligomerization CARD domains
[47]. Both receptors bind to the membrane of early endosomes and oligomerise upon ligand binding, leading to the activation of downstream signalling of NF-κB and MAPK pathways
[45], or the activation of autophagy independently of NF-κB
[48].
Both NOD1 and NOD2 are expressed in the skin, although NOD1 is mainly expressed by skin-homing immune cells
[49], while NOD2 expression is comparable in both immune cells and basal keratinocytes (A)
[50]. In human keratinocytes, NOD1 expression is induced by IFN-γ and was shown to activate IL-8 expression in response to
Pseudomonas aeruginosa [51]. NOD2 mediates beta-defensin-2 expression in keratinocytes subsequent to muramyl dipeptide exposure
[52].
4.2. NLRC5
NLRC5 is abundant in human skin with similar expression levels in keratinocytes to other cell types
[53][70]. NLRC5 has the highest homology to CIITA and also contains a nuclear localization sequence. The inclusion of NLRC5 in the NLRC family is due to the presence of a CARD-like domain
[54][71]. NLRC5 shuttles between the cytoplasm and nucleus and similar to CIITA induces the transcription of MHC class I genes in mouse and human cells
[55][56][57][72,73,74]. Like CIITA, NLRC5 also forms an enhanceosome and domain swapping experiments showed that the DD domain of both act as transcriptional activation domains
[58][75].
Besides its role in regulating antigen presentation, NLRC5 also plays a role in several cellular inflammatory reactions
[59][76]. In certain cell types, NLRC5 serves as a negative regulator of NF-κB activity by blocking the phosphorylation of IKKα and IKKβ
[60][61][77,78]. It was also reported to both promote and limit the antiviral type I interferon responses
[60][62][63][64][77,79,80,81]. NLRC5 was also demonstrated to cooperate with the NLRP3 inflammasome in response to bacterial infection
[65][82]. However, whether NLRC5 exerts any of these functions in the epidermis or in keratinocytes has not been studied so far.
4.3. Skin Diseases Associated with NLRC5 Functions
Skin Diseases Associated with NLRC5 Functions
Although NLRC5 functions have not been deeply studied in keratinocytes, its role in skin immune responses is supported by a genetic study. In a family, harbouring a pityriasis rubra pilaris (PRP)-causing gain-of-function
CARD14 mutation, the clinical manifestation was milder in a family member, who also carried an additional mutation of
NLRC5. This suggests a potential interplay of the two molecules in mediating the inflammatory response in keratinocytes
[66][83]. Interestingly, the epigenetic modifier Protein arginine methyltransferase 5 (PRMT5) downregulates NLRC5 expression in melanoma cells, leading to a decrease of MHC-I-mediated antigen presentation. Knockdown of PRMT5 promoted MHC-I accumulation at the cell surface of melanoma cells
[67][84]. NLRC5 is also highly expressed in keloids, fibrotic tumours in the skin that arise due to fibroblast hyperproliferation and increased expression of the extracellular matrix. Knockdown of
NLRC5 inhibits production of extracellular matrix components in keloid fibroblasts
[68][85].
5. The NLRP Subfamily
The NLRP subfamily has 14 members in humans, of which NLRP7, NLRP8, NLRP11 and NLRP13 have no orthologues in mice, while other members of the family have three (NLRP1, NLRP9), or seven (NLRP4) murine orthologues ()
[69][86]. NLRP proteins are characterized by the presence of an
N-terminal PYRIN domain, allowing the recruitment of the inflammasome-activating scaffold protein Apoptosis-associated speck-like protein containing a CARD (ASC)
[1]. Gene-expression of most NLRPs can be detected in immune cells and in keratinocytes
[15]. Besides protein expression in immune cells, NLRP1
[70][87], NLRP3
[71][88] and NLRP10
[14] are found in human skin samples, with other members being more predominantly expressed in other tissues.
Some members of the NLRP family (NLRP1, NLRP3, NLRP6, NLRP7 and NLRP12) form inflammasomes, leading to the activation of inflammatory caspases with subsequent IL-1β processing and release via inflammatory cell death, termed pyroptosis
[72][89]. Unlike professional immune cells, human keratinocytes do not need a priming signal to express inflammasome components, such as NLRP1, NLRP3, pro-IL-1β, ASC or pro-caspase-1
[15][70][73][15,87,90]. Moreover, the involvement of the NLRP subfamily to various skin diseases has been shown by association of genetic variants to inflammatory skin lesions, such as psoriasis, atopic dermatitis, and vitiligo as well as skin cancers
[74][75][76][77][78][79][91,92,93,94,95,96].
5.1. NLRP1
NLRP1 was the first described member of the NLRP subfamily to form inflammasome
[80][97]. There are three murine NLRP1 homologues to the gene:
Nlrp1a,
Nlrp1b,
Nlrp1c [1][81][1,98]. NLRP1 contains an
N-terminal PYRIN domain, a NACHT domain, LRRs, but also harbours a
C-terminal function-to-find domain (FIIND) and a CARD domain, through which it can directly activate caspase-1, albeit association with ASC enhances this activation
[82][99]. Interestingly, murine NLRP1 orthologues, Nlrp1a, -b and -c lack PYRIN domains
[83][100]. Activation of NLRP1 is unique among the NLRP family, as it undergoes auto-proteolysis within the FIIND and the resulting
N- and
C-terminal fragments remain non-covalently associated and auto-inhibited
[84][101]. Murine Nlrp1b was shown to be subsequently subjected to “functional degradation”, where the inhibitory
N-terminal domain is targeted for
N-end rule ubiquitination and proteasomal degradation, thus liberating and activating the
C-terminal fragment, which can form an inflammasome with capase-1
[85][102]. Human NLRP1 was initially described to be activated by muramyl dipeptide, while the murine protein is activated by
Bacillus anthracis lethal toxin
[86][87][88][103,104,105] and
Toxoplasma gondii [89][106]. More recently, NLRP1 was found to be activated in response to viral agonists, including dsRNA in keratinocytes, which binds directly to the NACHT-LRR region and induces ATP hydrolysis by the NACHT domain
[90][107]. Ultraviolet B irradiation (UVB) also induces activation of the NLRP1 inflammasome in human skin and keratinocytes, while murine keratinocytes fail to activate inflammasome upon UVB exposure
[70][73][87,90]. However, UVB irradiation does induce IL-1β release in murine skin, which is mediated by infiltrating professional immune cells (mainly dendritic cells) and not keratinocytes
[70][87]. These result show the poor conservation of the NLRP1 pathway between human and mouse skin
[91][108]. Interestingly, 3C proteases and dsRNA only activate human NLRP1, whereas Anthrax Lethal Factor metalloprotease induces cleavage of murine Nlrp1b but not the human form
[92][109].
T. gondii infection and Dpp8/9 inhibition with Talabostat (Val-boroPro) commonly activated both murine and human isoforms. Interestingly, Talabostat activates CARD8 in THP-1 cell-lines but triggers NLRP1 activation in keratinocytes, despite both cell types expressing both sensors
[91][108].
5.2. NLRP2
NLRP2 has a role in embryo development but its role in inflammasome activation is less clear. NLRP2 inhibits NF-κB activation but activates caspase-1 transcription
[93][115]. NLRP2 was reported to form inflammasomes in gingival epithelial cells but whether it is expressed or functional in epithelial skin is not elucidated
[94][116].
5.3. NLRP3
NLRP3 is the mostly extensively studied member of the NLRP subfamily and is predominantly expressed in immune cells. NLRP3 activation in immune cells requires a two-step process with a priming signal for transcriptional induction of signalling molecules, including NLRP3 itself, followed by a second, inflammasome activating signal. Both human and murine NLRP3 inflammasome activation can be initiated by numerous signals derived from cellular damage, such as ATP release
[95][118], potassium efflux
[96][119], reactive oxygen species, cathepsins and microcrystals
[97][120], but activation by microbial products has also been described
[98][99][121,122]. However, due to the diversity of these agonists, it is likely that NLRP3 does not bind directly to microbial PAMPs but instead detects cellular perturbation as part of a “Guard model”, similar to the detection of Rho GTPases by Pyrin after bacterial infection
[13].
Upon activation, NLRP3 undergoes a conformational change permitting NACHT domain-mediated oligomerisation. This aids recruitment of ASC to the PYRIN domain of NLRP3, and ASC further forms polymers that engages pro-caspase-1. Caspase-1 undergoes auto-cleavage and cleaves pro-IL-1β to its mature form
[100][123]. In addition, to this “canonical” inflammasome, NLRP3 can also engage in a second “non-canonical” inflammasome. In response to intracellular lipopolysaccharide, caspase-11 (in mouse, caspase-4 and -5 in humans) undergoes oligomerization and auto-activation. Caspase-11 cleaves Gasdermin D (GSDMD), which forms pores permitting the escape of mature IL-1β and IL-18
[101][124]. However, these pores upset the osmotic balance between the intra- and extracellular environments triggering pyroptosis. K
+ efflux as a result of these pores activates NLRP3 downstream of caspase-11
[102][125]. Caspase-1 also cleaves GSDMD, an event essential in pyroptosis, downstream of the canonical inflammasome
[103][126]. In macrophages, GSDMD cleavage by caspase-1 results in pyroptosis, but in human primary keratinocytes GSDMD is a poor substrate of caspase-1 and rather supports secretion of IL-1β, without pyroptosis
[104][127].
Although, NLRP1 is regarded as the principal inflammasome sensor in human keratinocytes
[70][79][91][87,96,108] expression of NLRP3 can also be detected in basal keratinocytes (A)
[105][106][128,129]. NLRP3 activation in human keratinocytes can be initiated by various signals
[105][106][128,129] but without a need for a priming signal. Human keratinocytes respond to viral RNA with caspase-1 activation and subsequent IL-1β and IL-18 release, which is dependent on NLRP3
[106][129]. Zhang and colleagues reported that soluble CD100 binds to PlxnB2 and activates NLRP3 in keratinocytes, leading to IL-1β and IL-18 release
[107][130]. UVB irradiation also indirectly causes NLRP3 inflammasome activation in keratinocytes by inducing cyclobutane pyrimidine dimer formation in the DNA, and the damaged DNA can induce NLRP3 inflammasome activation, leading to IL-1β release
[108][131]. Inflammasome activation is also indispensable for the normal wound healing processes, as demonstrated in mouse models, mainly due to macrophages and fibroblasts, not due to keratinocytes
[109][110][132,133]. However, overactivation of inflammasomes has an opposing effect, and inhibits normal wound healing
[111][112][134,135].
5.4. NLRP10
Both human and murine NLRP10 are highly expressed in the epidermis and contribute to cell-autonomous responses against invasive bacteria
[113][152]. Compared to other organs, murine epidermis was found to show the highest expression of Nlrp10 mRNA
[114][153]. Since NLRP10 lacks the prototypical
C-terminal leucine-rich repeats, it is thought to function as a signalling modifier. Indeed, NLRP10 positively regulates innate immune responses mediated by NOD1 upon
Shigella flexneri infection in both epithelial cells and dermal fibroblasts by modulating p38 MAPK and NF-κB signalling
[113][152]. Human NLRP10 inhibits ASC-mediated NF-κB activation and caspase-1 maturation of IL-1β
[115][154]. However, immune cells from
Nlrp10 knock-out mice respond normally to inflammasome activation
[116][117][155,156], suggesting a difference in function between the human and mouse proteins.
In mice, Nlrp10 has a bridging function between the innate and adaptive immune responses through DC activity. Against T-cell dependent antigens
Nlrp10-deficient mice show no efficient antigen specific immune responses due to impaired DC responses
[117][156], which also impairs the response to infection of the fungal pathogen
Candida albicans [116][155]. The lack of connection between the adaptive and innate immune system in the absence of Nlrp10 was further shown in other models.
Nlrp10 knock-out mice had significantly decreased inflammation in induced-contact hypersensitivity models and this was accompanied by a decreased infiltration of T cells. Mice with epidermal-specific knockout of Nlrp10 expression also displayed less inflammation but no loss of infiltrating T cell numbers
[118][157]. Interestingly, NLRP10 expression among other inflammasome regulators was strongly induced in differentiated human keratinocytes (A)
[14]. Moreover, GWAS studies also linked
NLRP10 genetic variants to atopic dermatitis
[119][78][55,95], a skin disease accompanied by abnormal differentiation and decreased barrier functions
[120][158]. These data strongly suggest a physiological role of NLRP10 in the skin, in addition to immune cells.
6. The NLRX Subfamily
The sole member of the NLRX family, NLRX1 contains a dissimilar and uncharacterised N-terminal effector domain compared to other NLRs. It also has an unusual C-terminus, which contains seven LRRs and a three-helix bundle [121][159]. Within the N-terminus NLRX1 contains a mitochondria-targeting sequence [122][123][124][160,161,162] and is involved in mitochondrial reactive oxygen species (ROS) formation [124][162]. Additionally, NLRX1 attenuates NF-κB and inflammasome signalling [124][125][162,163]. The regulatory effects of NLRX1 are highly cell type specific, which might be determined by the unique functional activity or metabolic profile of the given cell type [126][164]. NLRX1 is ubiquiteously expressed, including in keratinocytes, but its function in the skin is currently unknown.
(References would be added automatically after the entry is online)