The gut–liver axis plays important roles in both the maintenance of a healthy liver and the pathogenesis of liver diseases, where the gut microbiota acts as a major determinant of this relationship. Gut bacteria-derived metabolites and cellular components are key molecules that affect the function of the liver and modulate the pathology of liver diseases. Accumulating evidence showed that gut microbiota produces a myriad of molecules, including lipopolysaccharide, lipoteichoic acid, peptidoglycan, and DNA, as well as short-chain fatty acids, bile acids, trimethylamine, and indole derivatives. The translocation of these components to the liver exerts beneficial or pathogenic effects by interacting with liver immune cells. This is a bidirectional relationship.
- gut–liver axis,gut-microbiota,gut dysbiosis,liver disease,lipopolysaccharide,short-chain fatty acids
Please note: Below is an entry draft based on your previous paper, which is wrirren tightly around the entry title. Since it may not be very comprehensive, we kindly invite you to modify it (both title and content can be replaced) according to your extensive expertise. We believe this entry would be beneficial to generate more views for your work. In addition, no worry about the entry format, we will correct it and add references after the entry is online (you can also send a word file to us, and we will help you with submitting).
1. Introduction
The gut contains diverse microbial communities that have much more genetic material than the total human genome. The gut-residing bacteria have many health beneficial effects on the host by helping the gut to produce several metabolites, hormones, and peptides. The gut microbiota itself also produces several enzymes, metabolites, and cellular components that affect the health and diseases. The anatomy of the gut has a very close relationship with the liver through the portal vein. Therefore, the gut-derived components and immune signals are transferred to the liver, where they play a role in improving and maintaining the liver functions and healthy liver [1][2][1,2]. The liver is the largest gland with a remarkable function on the host by recruiting and activating the immune cells in response to gut-derived signals and components [3][4][3,4]. The liver produces primary bile acids that alter the gut microbial composition [5]. The interaction between the gut and the liver is bidirectional. Disruptions of this interaction result in the development of several liver diseases, including hepatic inflammation, alcoholic liver disease (ALD), non-alcoholic fatty liver disease (NAFLD), non-alcoholic steatohepatitis (NASH), fibrosis, cirrhosis, and hepatocellular carcinoma (HCC) [3][6][7][3,6,7]. Several therapeutic approaches have attempted to improve health by restoring the gut microbiota composition, production of metabolites, and modulation of immune signaling.
2. Gut Microbiota and Health
Gut-residing microbiota evolved with humans and became an essential organ in humans to influence health. In a healthy state, the gut microbiota has various beneficial effects in several ways (Figure 1). Four major phyla dominate in the GI: Bacteroidetes, Firmicutes, Proteobacteria, and Actinobacteria. Among them, Firmicutes and Bacteroidetes are the leading types followed by Proteobacteria, Actinobacteria, and a minor proportion of Verrucomicrobia and Fusobacteria phyla [8][9][8,9]. This bacterial colonization in the gut has a symbiotic relationship with the host via complex networks of interactions and crosstalk with each other. The interactions or crosstalk between the gut and liver is mainly through the gut microbiota-associated molecular patterns and their metabolic products that link the gut microbiota with other body organs system by acting as signaling molecules with immune regulatory functions.
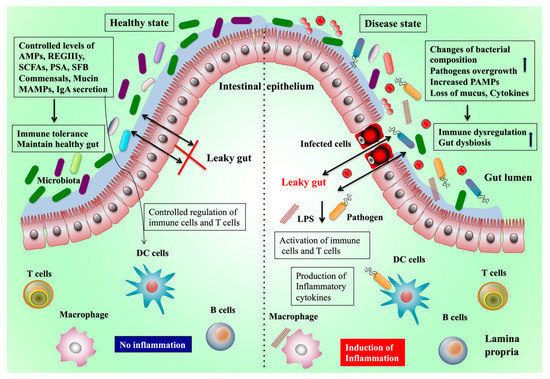
Figure 1.
Abbreviations used in the figure are: short-chain fatty acids (SCFA), antimicrobial peptides (AMPs), microbe-associated molecular patterns (MAMPs), pathogen-associated molecular patterns (PAMPs), inflammatory bowel diseases (IBD), regenerating islet-derived protein 3 γ (RegIIIγ), toll-like receptors (TLRs), segmented filamentous bacteria (SFB), and polysaccharide A (PSA).
3. Gut Microbiota and Diseases
Many factors, such as dietary products, antibiotic treatment, inflammatory products (LPS, flagella, LTA), and host physiological stress, have been shown to induce dysbiosis in gut microbiota composition. Dysbiosis impairs the functions of microbes and selectively influences the growth of pathobionts, which dysregulate the production of microbial products that induce the development of several diseases on local or neighbor organs, such as inflammatory bowel diseases (IBD), Clostridium difficile infection (CDI), obesity, metabolic syndrome, and diabetes. The decreased abundance of enteric bacterial diversity and the increased richness of Enterobacteriaceae have been strongly correlated with IBD [10][11][92,93], and the depletion of Enterobacteriaceae by the tungstate treatment ameliorates the severity of intestinal inflammation in mice with colitis [12][94]. The relatively higher abundance of Enterobacteriaceae and B. fragilis increased the levels of LPS that induce intestinal inflammation and colitis in mice through the suppression of Treg cells and the activation of TH1, and TH17 cells [13][95]. Faecalibacterium prausnitzii, an anti-inflammatory commensal bacterium, and its loss induces the reoccurrence of Crohn disease (CD), while its administration reduces inflammation by increasing anti-inflammatory cytokine (IL-10) and decreasing inflammatory cytokines (IL-12, IFN-γ) in a mouse model of colitis [14][96]. Clostridium difficile is a normal bacterial member in the gut, but it is also a pathobiont that reduces the intestinal epithelial cell integrity and induces inflammatory activity and cell death [15][97]. Clostridium difficile infection is often associated with antibiotic-mediated diarrhea that can be resolved by the dietary carbohydrate reduction [16][98].
Early findings indicated that the gut microbiota plays a role in the development of obesity in humans and animals. A lower diversity of gut microbes was observed in overweight and obese people [17][99]. Germ-free mice that received microbiota from obese subjects were heavier than the mice that received microbiota from healthy humans [18][100]. Perturbations in the composition of gut microbiota promote diet-induced obesity and metabolic diseases through different mechanisms, such as immune dysregulation, increased LPS production, altered energy, and gut hormone regulation [19][101]. The diet can also induce perturbations to the composition of gut microbiota and their functions. A study reported that mice fed a high-calorie Western diet showed a low number of Bacteroidetes and a high number of Firmicutes [20][102]. The high intake of sweeteners by humans or mice significantly altered the gut microbiota composition and induced glucose intolerance [21][103]. Rats fed sucralose exhibited an increased abundance of Bacteroides, Clostridia, and anaerobic bacteria in the gut [22][104]. Therefore, changes or loss of gut microbiota are associated with dysbiosis and are capable of inducing intestinal inflammation and other disorders that can be restored or ameliorated by supplementation with probiotics or prebiotics/dietary fibers [23][24][105,106]. In addition, studies suggest that alterations of the gut microbial composition extend its effects beyond the digestive system and can affect the functions of extra-intestinal organs, such as the liver. Figure 1 outlines how the gut microbiota contributes to pathogenic diseases and the mechanisms underlying the healthy and pathogenic state. This study discusses how the gut microbiota affects the liver function and promotes hepatic diseases via the gut–liver axis.
4. Gut–Liver Axis in Liver Diseases
Anatomically, the liver has a strong relationship with the gut where the gut microbes and their metabolites, nutrients, and gut-derived hormones contribute to the maintenance of healthy liver and liver metabolisms. The liver is not only a receiver of gut-derived products, but it also responds to the intestine by producing bile acids and IgA that affect the gut–liver axis [25][107]. Interestingly, liver-derived bile acid has been reported to induce changes in the gut microbiota composition [5]. Human and mouse liver contain gut-derived IgA-secreting cells that could mediate clearance of gut-derived antigens and protect the liver from pathogens, which reflects the strong connection between the gut and liver [26][108]. On the other hand, the bidirectional relationship between the gut and liver is normal in the healthy state, but during gut dysbiosis, a large number of gut bacteria and their derivatives, and microbial metabolites translocate to the liver, primarily through the portal vein, resulting in liver injury and the progression of liver diseases. The stage of liver disease truly relies on the severity of gut dysbiosis.
Studies have suggested the involvement of the gut microbiota in the development of non-alcoholic fatty liver disease (NAFLD), which is one of the most life-threatening liver diseases worldwide. Dysbiosis of the gut microbiota in the intestine of humans is associated with inflammation and impairments in mucosal immune function, which play a vital role in the pathogenesis of NAFLD [27][109]. Germ-free mice fed a high-fat diet (HFD) showed lower levels of lipids in the liver compared to HFD fed conventional mice [28][110]. In addition, germ-free mice received gut microbiota from hyperglycemia and insulinemia mice showed the development of NAFLD compared to the mice that received microbiota from normal mice [29][111]. Two bacterial species, Lachnospiraceae bacterium 609 and Barnesiella intestinihominis, were reported to be higher in the stool samples capable of inducing NAFLD, while B. vulgatus was found to be lower compared to the control group [29][111]. An increased abundance of Escherichia, Lactobacillus, Anaerobacter, and Streptococcus spp. was observed in NAFLD patients compared to healthy subjects [27][109]. Moreover, higher levels of TNF-α, IL-6, and IFN-γ, and a lower number of CD4+ and CD8+ cells were detected in NAFLD patients [27][109]. NAFLD can also develop into non-alcoholic steatohepatitis (NASH), which turns into fibrosis, cirrhosis, and hepatocellular carcinoma (HCC). Pathogen-free mice that received gut microbiota from HFD-fed mice showed gut vascular barrier (GVB) disruption and epidydimal adipose tissue enlargement through interference with the WNT/β-catenin signaling pathway in endothelial cells [2]. The increased GVB allows gut bacterial translocation to the liver, where they activate the parenchymal and non-parenchymal liver cells via pattern recognized receptors and others to develop various liver diseases [2][30][2,112]. Induction of intestinal inflammation increases gut-microbe-derived LPS in the portal circulation, resulting in hepatic inflammation and liver fibrosis, with the increased expression of collagen1, TIMP-1, TGF-β, PAI-1, and α- smooth muscle actin (SMA) in an experimental NASH mice model [31][113]. Liver fibrosis is a next stage of NASH. It can be induced by liver resident cells, especially hepatocytes, Kupffer cells (KCs), and hepatic stellate cells (HSCs). These cell types can express TLRs that are capable of recognizing microbe-derived ligands and mediate dangerous signals, resulting in liver injury.
5. Therapeutic Approaches
Several therapeutic approaches have been shown to restore the changes in gut microbiota composition and improve liver diseases, which include antibiotics treatment, prebiotics, probiotics supplementation, and fecal microbiota transplantation. Oral administration of antibiotics improved liver fibrosis [4] and alcohol-induced steatohepatitis [32][62]. On the other hand, a long-term approach may cause microbiome changes in the intestine of animals and humans. Probiotics are beneficial live microorganisms that have been shown to prevent liver diseases by reducing bacterial translocation and their derivatives and maintaining the gut barrier integrity [33][34][35][36][167,168,169,170]. L. rhamnosus R001 and L. acidophilus R0052 could ameliorate ALD in mice by reducing TLR4 expression and inflammatory cytokine (IL-6, IL-10, and TNF-α) production in mice [37][171]. Another study also showed that the development of ALD was prevented in mice treated with L. rhamnosus GG (LGG) by positively modulating bacterial composition to stimulate long-chain fatty acids (LCFAs) production, and increasing amino acid concentration in the intestine and liver of mice [38][172]. A combination of blueberry and probiotics has been reported to ameliorate NASH by increasing the level of peroxisome proliferator-activated receptor α (PPAR-α) and decreasing the level of sterol regulatory element-binding protein-1c (SREBP-1c), patatin-like phospholipase domain-containing protein 3 (PNPLA-3), and inflammatory cytokines (TNF-α, IL-6), Bcl-2 and caspase-3 in mice with NASH [39][173]. Clinical trials also confirmed the beneficial activity of probiotics against liver diseases, such as NAFLD/NASH and ALD in patients [40][41][174,175]. Prebiotics (inulin and pectin) are barely digestible food ingredients that have been reported to prevent hepatic injury, hepatic lipogenesis, and plasma triacylglycerol concentration by restoring Bacteroides in mice [42][176] and increasing SCFAs in humans [43][177]. Collectively, these studies provide evidence for the therapeutic potential of pre/probiotics on the amelioration of liver diseases, including NAFLD/NASH, ALD, and its related cirrhosis. There were no negative or adverse effects reported in clinical trials and mouse models using pre/probiotics.
Moreover, fecal microbiota transplantation (FMT) has attracted more attention for its ability to restore the gut microbiota and improve liver diseases. Alcohol-sensitive mice received FMT from alcohol-resistant donor mice that prevented the severity of ALD-induced liver injury, inflammation, steatosis and gut dysbiosis in vivo [42][176]. FMT improved rat behavior, hepatic encephalopathy (HE) grade and spatial learning capability in rat. In addition, FMT reduced intestinal permeability, intestinal mucosal barrier damage and systemic inflammation in rat treated with CCl4 [39][173]. Moreover, FMT in rat restored HE-induced losses of Claudin-1, Claudin-6 and Occludin in intestinal tissues of rat [44][178]. The liver disease indices were significantly improved within the first week after FMT therapy in patients with alcoholic liver disease [45][179]. Furthermore, FMT restored the gut microbiota composition, improved the metabolic pathways, bile secretion, carotenoid and pantothenate biosynthesis to normal levels in alcoholic liver disease patients. In a translational study, FMT from patients with alcoholic hepatitis induced severe inflammation, hepatic necrosis, intestinal permeability and bacterial translocation in germ-free humanized mice [46][180]. In addition, mice received from patients without alcoholic liver disease improved lesions in the liver, confirming the therapeutic roles of gut microbiota in alcoholic hepatitis.
In addition, herbal medicine (Sha-saiko-to) has potent effects and is used widely to treat patients with liver cirrhosis and HCC [47][181]. Herbal medicine has been shown to inhibit the activation of HSC cells, reduce hepatic lipid peroxidation, accumulation of extracellular matrix (ECM), expression of tissue inhibitor of metalloproteinases (TIMPs), and increase MMPs in rats [48][49][182,183]. A recent study reported that a Chinese herbal medicine (extract of Graptopetalum paragusyense) ameliorated dimethylnitrosamine (DMN)-induced hepatic inflammation and hepatic fibrosis via the suppression of TGF-β signaling in rats and rat HSC cells [50][184]. Overall, knowledge of the gut–liver axis has increased over the last decade through a series of microbiome studies that confirmed the vital role of the gut microbiota in chronic liver diseases.
6. Conclusions
The gut–liver axis plays a vital role in the etiology and pathogenesis of liver diseases. Gut-derived metabolites, cellular components, hormones, and others are translocated to the liver via the portal circulation, where they actively interact with immune cells and perform an inflammatory response, as well as induce the progression of several liver diseases. Among the gut-derived components, LPS is a key inflammatory molecule, which increasingly translocates to the liver during gut dysbiosis. Therefore, the TLR4/LPS signaling pathway is actively involved in the pathogenesis of liver diseases. In contrast, gut-derived SCFAs and BAs have beneficial effects that improve the liver functions. Current clinical and animal trials with different therapeutic strategies improve the present knowledge of the gut–liver axis, showing a favorable result that gives future hope to combat and ameliorate liver diseases.