Protein misfolding disorders are a group of diseases characterized by supra-physiologic accumulation and aggregation of pathogenic proteoforms resulting from improper protein folding and/or insufficiency in clearance mechanisms. Although these processes have been historically linked to neurodegenerative disorders, such as Alzheimer’s disease, evidence linking protein misfolding to other pathologies continues to emerge. Indeed, the deposition of toxic protein aggregates in the form of oligomers or large amyloid fibrils has been linked to type 2 diabetes, various types of cancer, and, in more recent years, to preeclampsia, a life-threatening pregnancy-specific disorder. While extensive physiological mechanisms are in place to maintain proteostasis, processes, such as aging, genetic factors, or environmental stress in the form of hypoxia, nutrient deprivation or xenobiotic exposures can induce failure in these systems. As such, pregnancy, a natural physical state that already places the maternal body under significant physiological stress, creates an environment with a lower threshold for aberrant aggregation.
- protein misfolding
- preeclampsia
- amyloid-β
- Alzheimer’s disease
- tau
- pregnancy
1. Introduction
2. The Central Hypothesis of PE Pathophysiology and Current Gaps
While substantial progress has been made over the years to understand the pathophysiological mechanisms associated with PE, significant controversy remains. A chief challenge is that PE is not a single disease with a single causative pathway but rather a collection of highly heterogeneous clinical phenotypes with non-specific molecular and pathologic findings that often overlap with other pregnancy disorders [17,18][11][12]. Clinically, PE manifests as new-onset hypertension (systolic blood pressure ≥ 140 mmHg, diastolic blood pressure ≥ 90 mmHg) after 20 weeks of gestation with or without proteinuria [17][11]. This definition is a recent revision of the traditional clinical classification criteria for PE, which was based on the combination of hypertension and proteinuria [19,20][13][14]. The definition has been broadened to encompass cases without proteinuria but with evidence of maternal end-organ damage in the form of hemolysis, elevated liver enzymes and low platelet count syndrome (HELLP); renal damage; pulmonary edema; visual disturbances; persistent headache; stroke; and seizures (eclampsia) [19][13]. The current diagnostic and clinical guidelines distinguish between PE with and without severe features (sPE) based on the occurrence of any of these severe maternal symptoms or a systolic blood pressure over 160 mmHg and diastolic blood pressure above 90 mmHg [19][13]. As a result of this revision, an increased number of women with milder symptoms and a lower probability of adverse outcomes are now diagnosed with PE [21][15]. On the other hand, the clinical phenotype can also be subclassified based on the time of disease onset. PE arising before 34 weeks of gestation classifies as early-onset PE (EOPE) while PE arising after 34 weeks of gestation classifies as late-onset PE (LOPE). LOPE and EOPE have been shown to have distinct pathophysiological mechanisms and varying levels of disease severity [17][11]. It is widely accepted that the placenta, a transient but indispensable organ for pregnancy maintenance and fetal development, is the initial site where dysfunction arises. Indeed, the central hypothesis of PE pathogenesis supports a two-stage model with a defective invasion of extravillous trophoblasts cells (EVTs) and incomplete remodeling of spiral arteries at the maternal–fetal interface during placentation as the inciting factor for disease (Stage 1) [23,24,25][16][17][18]. In normal pregnancy, spiral arteries, which connect the placenta to the maternal blood supply, undergo extensive transformation. These blood vessels must widen to accommodate the increase in blood flow necessary to support a growing fetus [26][19]. This remodeling process has been shown to be mediated by the invasion of EVTs from the anchoring villi to the maternal decidua and deeper into the myometrium (interstitial trophoblasts). A subpopulation of the migrating EVTs also invades the wall and lumen of spiral arteries, where they replace endothelial cells (as intramural and endovascular trophoblasts, respectively) [26,27,28,29,30][19][20][21][22][23]. Despite well-established evidence of this pathophysiologic process occurring in PE, the shallow trophoblast invasion characterizing Stage 1 is not unique to PE. The failure of spiral artery transformation has also been described in preterm birth, fetal growth restriction (FGR), placental accreta spectrum disorders, and spontaneous miscarriage with varying degrees of changes and severity in placental phenotype [32,34,36,37,38,39][24][25][26][27][28][29]. Furthermore, although signs of abnormal placentation are commonly observed, they are not observed in all affected women, nor are they always sufficient to trigger clinical symptoms when present [29,40][22][30]. Intriguingly, the endothelial dysfunction resulting from placental malperfusion causes an imbalance in angiogenic markers, such as soluble fms-like tyrosine kinase-1 (sFlt-1) andsoluble endoglin (sEng), which increase; and VEGF which decreases [41,42,43][31][32][33]. However, there is evidence that these changes in levels of angiogenic factors can be detected as early as 7 weeks of gestation, before the deep invasion of EVTs occurs, which suggests that vascular remodeling dysfunction may occur independently of trophoblast invasion [17,33,44,45][11][34][35][36]. A multitude of environmental, genetic, fetal, and maternal risk factors that modulate PE risk and influence disease severity also exist. One striking example is the relationship between chronic kidney disease (CKD) and PE. On one hand, PE onset is associated with the development of acute end-organ damage involving vascular damage, glomerular endotheliosis, and chronic kidney damage, with a three–fifteen-fold increased relative risk of subsequent end-stage renal disease [53,54][37][38]. On the other hand, women who have pre-existing CKD are ten times more likely to develop PE compared to women without CKD [55,56][39][40]. Even more intriguing, women with a single kidney are three times more likely to develop PE than women with two kidneys, even in the setting of a normal glomerular filtration rate [57][41]. This epidemiological observation has also been demonstrated in vivo in a mouse model that exhibits a PE-like phenotype following unilateral nephrectomy [58][42]. The spectrum of potential placental, fetal, and maternal pathophysiological processes and risk factors involved in PE create considerable ambiguity in discriminating cause from effect. Hence, a multifactorial etiology of PE that involves abnormal placentation, immune maladaptation mechanisms, cardiovascular and renal dysfunction, as well as multisystemic disruptions in proteostasis cannot be dismissed (Figure 1).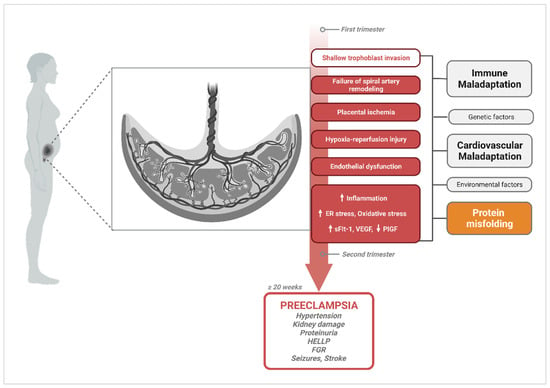
3. Protein Misfolding in PE
3.1. General Protein Misfolding and Aggregation Mechanisms
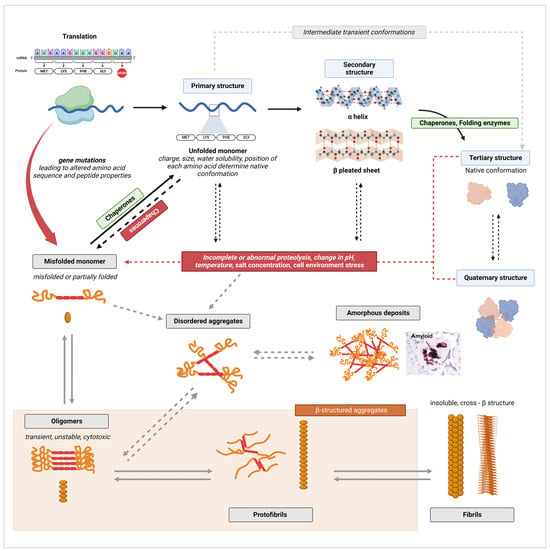
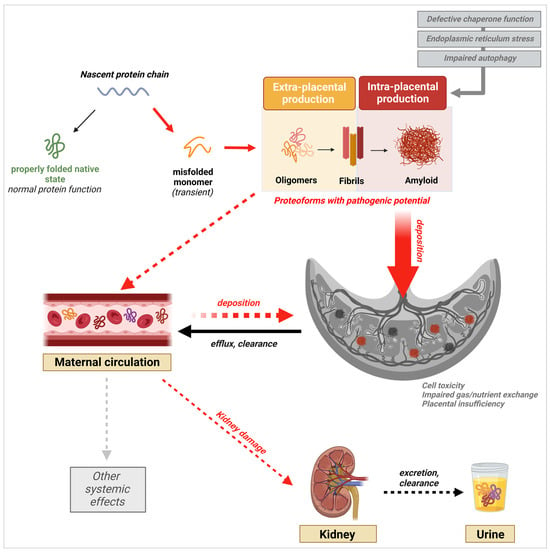
3.2. Excretion of Misfolded Proteins in the Urine
3.3. Presence of Misfolded Proteins in the Maternal Circulation
Kalkunte et al. demonstrated the presence of aggregated proteoforms of transthyretin (TTR) in the serum of women with PE, providing additional supporting evidence for the protein misfolding hypothesis of PE pathophysiology [11][67]. They also showed that the intraperitoneal injection of PE serum or TTR directly extracted from PE serum induced a PE-like syndrome characterized by hypertension, glomerular endotheliosis, fetal growth restriction, proteinuria, and increased sFlt-1 and sEng in IL-10-/- mice compared to mice injected with serum from normal pregnancies [11][67]. The PE-like phenotype was rescued when PE serum was co-administered with exogenous TTR in its native form or when TTR was depleted from PE serum [11][67]. These findings highly suggest that the presence of these abnormal TTR aggregates is causative of PE and not a result of any preceding dysfunction. Using a cell-based in vitro model for the detection of protein aggregates, Cheng et al. also reported the presence of Amyloid-β in addition to TTR in the serum of women with EOPE and LOPE [15][68]. Using the same technique, Jash et al. confirmed that aggregated forms of the cis stereo-isoform of P-tau231 (phosphorylated at threonine residue 231), a protein isoform linked to pathogenicity in AD, are present in the serum of women with EOPE and LOPE but not the gestational-age-matched controls. The presence of multiple proteins that have been proven to be pathogenic in AD in PE reinforces the idea that the aggregation and accumulation of these proteoforms in PE serum is not harmless.3.4. Placental Deposition of Extracellular Protein Aggregates
A group has previously reported the presence of amyloid-like deposits of SERPINA1, APP, and Aβ in the placentas of women with sPE [12,13][64][65]. Detection of SERPINA1 aggregates using ATZ11, an antibody specific for oligomeric conformations of the protein, revealed increased placental deposition of misfolded SERPINA1 complexes in the stroma, endothelium, and fetal blood vessels of placentas from sPE pregnancies compared to gestational-age-matched controls [13][64]. Furthermore, these deposits were predominantly localized in the endothelium. Given the central role endothelial damage plays in PE pathophysiology, and the well-documented toxicity of oligomeric protein conformations, the perivascular localization of these deposits could be an indication that they contribute to endothelial dysfunction [86,95][62][69]. Abnormal proteolytic cleavage of APP has been established as the mechanism for Aβ formation, aggregation, and accumulation in the AD brain [59,87,96][43][63][70]. Similarly, hyperphosphorylation of tau, leading to the aberrant accumulation of pathogenic conformations of the protein, occurs [96,97][70][71]. Together, the deposition of toxic oligomers and large extracellular fibrillary plaques of Aβ with neurofibrillary tangles made of hyperphosphorylated tau are the two principal hallmarks of AD pathology. Along with the detection of misfolded SERPINA1, a group identified the presence of APP aggregates in the urine and placentas of women with sPE. Immunostaining of the placental tissue with ALZ90, an antibody specific for the Aβ fragment of APP, showed a positive signal for amyloid-like plaques of Aβ within areas of fibrinoid deposition [12][65]. This phenotype was accompanied by significant upregulation of β-secretases, the enzymes that are responsible for the abnormal cleavage of APP into Aβ, in the trophoblast layer of placental villi. These observations led us to propose that the placental or systemic clearance of misfolded proteins might be deficient in PE, leading to a supraphysiologic load of these species in the urine. Following the detection of aggregated proteoforms of TTR in PE serum, extensive studies have been conducted to understand their role in PE-associated placental pathology. Studies by Kalkunte et al. and Tong et al. revealed TTR deposition in the villous stroma and extravillous trophoblast cell layer in the human placental villi section [11,99][67][72]. The regions with positive TTR staining matched Thioflavin-S-positive regions, confirming that the TTR placental deposits in question were aggregated forms of the protein [11][67]. Previous work by Kalkunte et al. demonstrated that PE serum containing TTR aggregates also caused the disruption of endovascular activity and crosstalk between endothelial cells and trophoblasts. Although these latest research findings shed light on the placental metabolism of aggregated proteins, like Aβ, cis P-tau, and TTR, and offer a potential source for the aggregated proteoforms detected in serum, the conditions in which specific proteins are formed are still unclear. It also appears that pathogenic forms from different proteins may impact placental function at the same time, with some being more predominant than others. A deeper look into the function and characteristics of each of these proteins may help explain their presence and effects in PE.4. PE-Associated Misfoldome
4.1. SERPINA1/α-1-Antitrypsin (A1AT)
Serine protease inhibitors, also known as serpins, are a superfamily of proteins involved in the regulation of proteolytic enzymes throughout the body [100][73]. Serpins, especially SERPINA1, have been involved in a multitude of pathogenic processes, leading to a group of disorders recognized as serpinopathies. Notably, genetic mutations in SERPINA1 lead to severe and uncontrolled lung damage due to failure in the inhibition of neutrophil elastase, an abundant protease in the lung. In the liver, the main site of the synthesis of SERPINA1, defective SERPINA1 molecules accumulate and aggregate into higher-order polymers that induce fibrosis and cirrhosis [101][74]. SERPINA1 is also an acute-phase reactant that is upregulated during inflammation, infection, and pregnancy [101][74]. Thus, a role for the protein in PE pathophysiology is not unlikely. For example, the protein has well-documented anti-inflammatory properties that are independent of its proteolytic function [108][75]. The increased placental and systemic inflammation, which are characteristic pathophysiological processes in PE, could potentially drive increased SERPINA1 expression as a protective mechanism [92,109][76][77]. On the other hand, Twina et al. showed that sPE is associated with lower circulating levels of SERPINA1, in which case dysregulation in SERPINA1 levels could be the cause for increased inflammation [110][78]. It has also been proposed that SERPINA1 protects against the development of PE through the suppression of oxidative stress both in vitro and in vivo [111,112][79][80]. An important question is whether any of the known mutations that induce SERPINA1 oligomerization and deficiency exist in PE. Nagarajappa et al. conducted a cross-sectional study of 200 preeclamptic women to characterize the frequency of PiS and PiZ mutations, the two dominant mutations in the SERPINA1 gene that cause A1AT deficiency [116][81]. Results showed that these two variants were non-existent in the study cohort; however, the presence of other unidentified pathogenic mutations cannot be excluded [117][82]. The origin of SERPINA1 dysregulation and misfolding in PE is still unclear. Supposing the dysregulation results from established pathology as a compensatory mechanism instead of triggering it, there is still a high potential for a double-whammy effect due to the protein’s propensity to aggregate into toxic oligomers. The presence of highly amyloidogenic SERPINA1 fragments, such as the FVFLM peptide in the urine of women with PE, supports that excessive synthesis and release of toxic SERPINA1 proteoforms in the circulation either by the placenta or other organs occurs. The resulting urinary excretion of these peptides could be a clearance mechanism that helps remove these toxic species from the body [13,93][64][83].4.2. Aβ and Its Precursor APP
Despite ample evidence of the presence of misfolded Aβ and APP in PE serum, placenta, and urine, mechanistic studies on their effects in PE are sparse [12,15,94][65][68][84]. However, the aggregation of Aβ, which is a pathognomonic feature of AD pathology, is an uncanny similarity that can be exploited to better understand the mechanisms at play in PE. Thus, based on knowledge gathered from studies of Aβ pathology in AD, it can make inferences on what processes involving Aβ aggregation might be at play in PE. In AD, the amyloid cascade hypothesis is the classic view of pathogenesis [96,119][70][85]. According to this hypothesis, the deposition of Aβ aggregates drives neurotoxicity and the onset of dementia in AD [119][85]. Aβ monomers are a cleavage product resulting from the non-canonical processing of APP by β- and γ- secretases enzymes (BACE1, BACE2, PS1, PS2) instead of α-secretases (ADAM10) [120,121][86][87].4.3. Tau
Alongside Aβ, tau aggregation is a major driver and hallmark of AD symptomatology. Tau is a microtubule-associated protein that aids in the assembly and stabilization of microtubule filaments. In the nervous system, these functions are critical for neuronal maturation and axonal transport [124][88]. Critically, the protein’s function is regulated via multiple post-translational modifications, the most common of which is phosphorylation at various residues [125][89]. Phosphorylation of tau induces its dissociation from microtubules, causing alterations in cytoarchitecture and poor microtubule stability [124,125][88][89]. In AD, and other tau-associated proteinopathies, hyperphosphorylation of tau at specific sites is known to induce neurotoxic effects, including self-aggregation into pathogenic helical filaments that make up neurofibrillary tangles and toxic intracellular inclusions [125][89].4.4. TTR
TTR is an essential carrier protein that transports thyroxine and retinol-binding protein (RBP) bound to retinol (Vitamin A) [126][90]. It is primarily synthesized by the liver, choroid plexus, and retina before being secreted into the circulation and cerebrospinal fluid (CSF) [126][90]. Studies have shown that placental villous trophoblasts also synthesize TTR and that, in PE, an increased amount of placental extracellular vesicles carrying aggregated TTR as a cargo were released into maternal circulation [99,127][72][91]. One essential function of TTR that could also contribute to PE pathophysiology is its role in angiogenesis and maintaining blood–brain barrier integrity in AD [129,130][92][93]. Nunes et al. showed that V30M, an amyloidogenic hereditary variant of TTR, causes the downregulation of pro-angiogenic factor expression, induces apoptosis, and reduces cell migration in endothelial cells [130][93]. Analogous effects of TTR aggregates on placental vasculature, including the impairment of capillary tube formation and induction of the anti-angiogenic factors sFlt-1 and sEng, have been described by Kalkunte and al [11][67]. TTR mutations like V30M or age-driven misfolding and aggregation of wild-type TTR are involved in familial amyloid polyneuropathy (FAP) and senile systemic amyloidosis (SSR), respectively [129][92]. Both disorders are characterized by the deposition of TTR fibrils in the peripheral nervous system in the case of FAP and, primarily, in cardiac tissue for SSR. The pathophysiology of these disorders mimics that of other proteinopathies, including the characteristic tissue and cellular damage inflicted by toxic oligomeric intermediates [131][94].5. Driving Mechanisms of Protein Misfolding in PE
5.1. ER Stress and Hyperactivation of the Unfolded Protein Response (UPR)
ER stress is a well-defined feature of PE [23,35,60][16][44][95]. It is also a leading pathophysiologic process in protein conformational disorders [137][96]. The ER is a focal site of protein synthesis and maturation. Following mRNA translation by ribosomes in the cytoplasm, peptides are translocated to the ER lumen, where they undergo necessary post-translational modifications, folding, and assembly into their functional conformation. To carry out these functions properly, the microenvironment in the ER lumen must maintain a high concentration of Ca2+ and a high oxidative capacity [60][44]. In PE, chronic low oxygen tension in the placenta, as a result of impaired uteroplacental perfusion, disrupts the delicate balance of the ER milieu, therefore inhibiting proper function [60,109][44][77]. This state of ER stress leads to the accumulation of unfolded proteins, which are more prone to misfold and aggregate, as the ER functional capacity is diminished [138][97]. Common ER stress markers, such as ATF6, GRP78 (BiP/HSPA5), Ire1α, CHOP, PERK, and eIF2α, are upregulated in the PE placenta [109,134,138,139,140,141,142,143][77][97][98][99][100][101][102][103]. While UPR activation normally acts as a protective mechanism, an excessive and prolonged UPR, as would be the case during chronic tissue hypoxia, is self-damaging. Ultimately, cellular response mechanisms can be overwhelmed, leading to a collapse of protective systems and dooming cells for apoptosis.5.2. Failure of the Protein Folding Machinery: The Role of Chaperones
Transcriptional upregulation of chaperones and folding enzymes is one of the compensatory mechanisms that are enacted by the UPR response. Under normal physiologic conditions, chaperones are molecules that aid other proteins in acquiring their natural conformation. They also have the capacity to recognize and bind to misfolded proteins, hence inhibiting or undoing their aggregation; and the capacity to unfold and refold improperly folded polypeptides, or tag them for degradation [61][45]. ER stress is not the only inducer of chaperone activity. For example, heat shock proteins (HSPs) are a ubiquitously expressed family of chaperones that are upregulated in response to a variety of stressors that include heat, oxidative stress, sterile inflammation, and viral infection [148,149][104][105]. Members of the HSP70 family, in particular, are notoriously involved in the regulation of decidual and placental cell function from placentation to the end of pregnancy [150][106]. Their upregulation in PE not only serves as a marker of UPR activation but also indicates that their overactivation could be driving pathological processes. If misfolded proteins are present in excess, increased expression of HSP70 isoforms like GRP78 is necessary for re-folding and disaggregating already tangled proteins. On the other hand, when bound to misfolded proteins, they are no longer available to perform other functions that are critical to maintaining cell integrity. This has been proposed as another possible mechanism driving the toxicity of protein aggregates like polyglutamine, an amyloidogenic protein involved in Huntington’s disease pathogenesis [151][107]. In PE-associated misfolding, pregnancy zone protein (PZP) has also emerged as an important player. In contrast with GRP78, which is an ER-associated intracellular chaperone, PZP has recently been characterized as an extracellular chaperone that can inhibit the aggregation of Aβ [135][108]. PZP is a dimeric homolog of the alpha-2-macroglobulin (α2M) that is found in high concentrations in the serum of pregnant women while levels are characteristically low in non-pregnant women and men [152,153,154][109][110][111]. With a 71% degree of homology between the two protein sequences, it has been proposed that they may perform similar functions [152,155][109][112]. Plasminogen activator inhibitor type 2 (PAI-2), another extracellular chaperone, has been linked to PE-associated protein aggregation [136][113]. Similarly to PZP, PAI-2, also known as SERPIN2B, is normally upregulated in pregnancy [166][114]. PAI-2 plays an important role in inflammation through the regulation of T-helper-cell activation [167][115]. The protein is expressed by activated macrophages but also endothelial cells and placental trophoblasts [166][114]. With data indicating an association between reduced PAI-2 mRNA and protein levels, increased Th1 activation, placental insufficiency, and endothelial dysfunction in PE, PAI-2 has been proposed as a potential contributor to pathogenesis in pregnancy. [52,167,168,169][115][116][117][118]. The serpin can also act as a holdase, a type of chaperone protein that can passively bind to prefibrillar protein species, preventing them from further aggregation [166,170,171][114][119][120].5.3. Failure in Clearance Mechanisms: Defective Autophagolysosomal Processing
The third level at which disruptions in proteostasis mechanisms can occur is after all attempts at slowing the production of misfolded proteins and enhancing folding mechanisms have failed. At that time, misfolded proteins that exceed the UPR’s corrective capacity are targeted for degradation. Several studies and reviews have examined defects in autophagy as a potential root cause of protein aggregate accumulation in PE [14,122,134,181,182,183,184][98][121][122][123][124][125][126]. Autophagy is one of two processes through which cells clear defective proteinssuch as aggregated proteins, and, damaged intracellular components like organelles [63][47]. This process is mediated by a degradative enzyme in the lysosomes after the organelle fuses with autophagosomes carrying damaged cellular components (macroautophagy) or after the direct intake of individual unfolded proteins that were preemptively tagged (by ubiquitination) for degradation by chaperones (microautophagy) [185][127]. The second mechanism through which intracellular proteolysis occurs is through the ubiquitin–proteasome pathway. Upon ubiquitination, proteins may diverge to the proteasome, a large protein complex that can hydrolyze and degrade proteins [186][128]. Nakashima et al. postulated a decade ago that autophagy induction is critical to protecting placental trophoblast cells from hypoxia-induced disruptions. They showed that relative to autophagy-competent cells, autophagy-deficient trophoblast cells exhibited poor invasion and decreased ability to induce vascular remodeling in response to hypoxia, suggesting that defects in autophagy may contribute to poor vascular remodeling [184,187][126][129]. Cheng et al. showed that both the blockade of autophagosome–lysosome fusion with chloroquine and that of proteasome activity via MG132 treatment cause the accumulation of TTR aggregates in PHTs [134,188][98][130]. Moreover, there was an irregular placental accumulation of ubiquitinated proteins in the setting of EOPE-associated ER stress and UPR. These results, while preliminary, support that the impairment of autophagy, but also of ubiquitination-mediated protein degradation processes, may play a role in PE-associated protein aggregation [134][98].6. New Prediction, Diagnostic, and Treatment Approaches Based on PE-Associated Protein Misfolding
6.1. Early Prediction and Diagnosis
To date, accurate and timely PE diagnosis continues to pose a major challenge in clinical practice. The recent exclusion of proteinuria as a compulsory diagnostic criterion was favorable for the detection of non-proteinuric PE cases but left the presence of elevated blood pressure as the only sign that can be used to initiate further [19,197][13][131]. Based on current clinical diagnostic guidelines, the syndrome can only be detected when overt symptoms are already present. Methods that have been developed in AD can inform us of the potential of misfolded protein detection to accurately predict and diagnose proteinopathies. For example, plasma and CSF assays assessing Aβ42/Aβ40 and tau/P-tau levels among other biomarkers are highly successful at identifying the presence of AD pathology prior to the threshold for amyloid positivity on PET scans and the onset of cognitive decline and at predicting disease trajectory [199,200,201][132][133][134]. The biomarker assays perform even better when combined with genetic risk score [202][135]. APOE-ε4, one of three alleles of the APOE gene, is well-established as the greatest genetic risk factor for AD. Adding APOE-ε4 status alone to prediction models increases AUC and, thus, predictive value [202,203][135][136]. A novel blood-based assay that can detect the presence of misfolded Aβ, TTR, or P-tau in the maternal circulation has been proposed by Cheng et al. [15][68]. This method uses an autophagy-deficient trophoblast cell model to establish whether misfolded proteins are present in serum. The exposure of these specific autophagy-deficient human trophoblasts (ADTs) to serum from patients with PE, AD, and mild cognitive impairment leads to the intracellular accumulation of protein aggregates made of TTR and Aβ in PE cases, with the addition of α-synuclein in AD and MCI cases [15][68]. The Congo red dot (CRD) test represents a non-invasive and low-cost alternative to other methods of assessing protein misfolding load associated with PE. The clinical applicability of the CRD test, which is based on the detection of urine congophilia, has been examined in fourteen studies, including one randomized control trial and one meta-analysis ([90,91,204,205,206,207,208,209,210,211,212,213,214,215][137][138][139][140][141][142][143][144][145][146][147][148][149][150]). On the other hand, a few studies report low test performance. Results from Sammar et al. showed that the CRD test can diagnose PE with high accuracy in women presenting with symptoms; however, that the test is less effective at predicting PE in the first trimester of pregnancy when used alone [208][143]. The predictive value of the test was improved when %CRR was used in combination with other markers of high disease risk, such as previous PE diagnosis, black race, body mass index, and mean arterial pressure [208][143].6.2. Treatment
Other than the emergent delivery of the fetus and placenta, which comes with its own set of iatrogenic complications when conducted preterm, there is no effective therapy for PE. Novel strategies that can be employed to reduce the accumulation of misfolded proteins and prevent their toxic effects include the inhibition of ER stress, stabilization of chaperone function, or restoration of autophagic balance. For example, the stabilization of TTR’s tetrameric structure is currently being explored as a therapeutic mechanism for the treatment of FAP and AD. Dissociation of the tetramer into monomers potentiates TTR’s aggregation into fibrils in FAP and compromises TTR’s ability to sequester unstable Aβ conformations in AD [129,132,216,217][92][151][152][153]. ER stress modulators, autophagy-enhancing drugs, gene therapy, and anti-amyloid immunotherapy are also being examined as therapeutic approaches in the treatment of AD [146,220,221,222][154][155][156][157]. In recent years, sildenafil, a known inducer of autophagy, has been associated with the alleviation of cognitive impairment in pre-clinical models and decreased incidence of AD in human studies and, thus, has been proposed as a candidate drug that could be repurposed for AD treatment [223,224][158][159]. The potential of sildenafil use for the treatment of AD has not yet been verified as evidence from a recent study does not support previous claims [225][160].7. Long-Term Implications of Pregnancy-Related Protein Misfolding Disease on Maternal and Fetal Health
As the field progresses in its understanding of PE-associated disease processes, the more complex the etiology of the disorder seems to become. Current evidence seems to point to a multisystem multifactorial pathological process, which extends beyond placental dysfunction. Gaining a better understanding of protein misfolding mechanisms and how they may contribute to systemic dysfunction in PE is crucial to advancing the ability to care for mothers and their babies during pregnancy. On the other hand, it also gives us a window into their future health. It is well-established that having PE during pregnancy greatly increases the risk of cardiovascular disease later in life for both mothers and their offspring [237,238,239,240][161][162][163][164]. Accumulating epidemiological reports also suggest a link between hypertensive disease during pregnancy and long-term risk of vascular dementia and AD [241,242][165][166]. While the discovery of significant protein misfolding pathology in PE opens many new avenues for investigation, diagnosis, and treatment, many questions are left to be answered. The central questions that persist and still require attention include whether protein aggregation instigates PE or is a consequence of it; the reasons behind the selective detection of certain proteins in specific tissues (serum vs. placenta vs. urine); and the systemic factors that may affect the accumulation and deposition of misfolded proteins in pregnancy. The possibility that proteostasis disruption might occur systemically rather than being confined at the placental level carries significant implications for both immediate and long-term maternal and fetal health.References
- Labbadia, J.; Morimoto, R.I. The Biology of Proteostasis in Aging and Disease. Annu. Rev. Biochem. 2015, 84, 435–464.
- Bradbury, J. Chaperones: Keeping a Close Eye on Protein Folding. Lancet 2003, 361, 1194–1195.
- Chaudhuri, T.K.; Paul, S. Protein-Misfolding Diseases and Chaperone-Based Therapeutic Approaches. FEBS J. 2006, 273, 1331–1349.
- Powers, E.T.; Morimoto, R.I.; Dillin, A.; Kelly, J.W.; Balch, W.E. Biological and Chemical Approaches to Diseases of Proteostasis Deficiency. Annu. Rev. Biochem. 2009, 78, 959–991.
- Williams, D. Pregnancy: A Stress Test for Life. Curr. Opin. Obstet. Gynecol. 2003, 15, 465.
- Williams, D.J. Physiological Changes of Normal Pregnancy. In Oxford Textbook of Medicine; Warrell, D.A., Cox, T.M., Firth, J.D., Eds.; Oxford University Press: Oxford, UK, 2010; ISBN 978-0-19-920485-4.
- Cunningham, F.G.; Leveno, K.J.; Bloom, S.L.; Dashe, J.S.; Hoffman, B.L.; Casey, B.M.; Spong, C.Y. Maternal Physiology. In Williams Obstetrics, 25e; McGraw-Hill Education: New York, NY, USA, 2018.
- Soma-Pillay, P.; Catherine, N.-P.; Tolppanen, H.; Mebazaa, A.; Tolppanen, H.; Mebazaa, A. Physiological Changes in Pregnancy. Cardiovasc. J. Afr. 2016, 27, 89–94.
- Kalhan, S.C. Protein Metabolism in Pregnancy. Am. J. Clin. Nutr. 2000, 71, 1249S–1255S.
- Duggleby, S.L.; Jackson, A.A. Protein, Amino Acid and Nitrogen Metabolism during Pregnancy: How Might the Mother Meet the Needs of Her Fetus? Curr. Opin. Clin. Nutr. Metab. Care 2002, 5, 503.
- Huppertz, B. Placental Origins of Preeclampsia. Hypertension 2008, 51, 970–975.
- Burton, G.J.; Redman, C.W.; Roberts, J.M.; Moffett, A. Pre-Eclampsia: Pathophysiology and Clinical Implications. BMJ 2019, 366, l2381.
- Gestational Hypertension and Preeclampsia: ACOG Practice Bulletin, Number 222. Obstet. Gynecol. 2020, 135, e237.
- Copel, J.A.; Platt, L.D.; Hobbins, J.C.; Afshar, Y.; Grechukhina, O.; Mallampati, D.; Bromley, B.; Caughey, A.B.; Grobman, W.; Han, C.S.; et al. Gottesfeld-Hohler Memorial Foundation Risk Assessment for Early-Onset Preeclampsia in the United States: Think Tank Summary. Obstet. Gynecol. 2020, 135, 36.
- Reddy, M.; Fenn, S.; Rolnik, D.L.; Mol, B.W.; da Silva Costa, F.; Wallace, E.M.; Palmer, K.R. The Impact of the Definition of Preeclampsia on Disease Diagnosis and Outcomes: A Retrospective Cohort Study. Am. J. Obstet. Gynecol. 2021, 224, 217.e1–217.e11.
- Chaiworapongsa, T.; Chaemsaithong, P.; Yeo, L.; Romero, R. Pre-Eclampsia Part 1: Current Understanding of Its Pathophysiology. Nat. Rev. Nephrol. 2014, 10, 466.
- Roberts, J.M.; Hubel, C.A. The Two Stage Model of Preeclampsia: Variations on the Theme. Placenta 2009, 30, S32–S37.
- Redman, C.W.G.; Sargent, I.L. REVIEW ARTICLE: Immunology of Pre-Eclampsia. Am. J. Reprod. Immunol. 2010, 63, 534–543.
- Brosens, I.; Robertson, W.B.; Dixon, H.G. The Physiological Response of the Vessels of the Placental Bed to Normal Pregnancy. J. Pathol. Bacteriol. 1967, 93, 569–579.
- Pijnenborg, R.; Bland, J.M.; Robertson, W.B.; Brosens, I. Uteroplacental Arterial Changes Related to Interstitial Trophoblast Migration in Early Human Pregnancy. Placenta 1983, 4, 397–413.
- Pijnenborg, R.; Vercruysse, L.; Hanssens, M. The Uterine Spiral Arteries in Human Pregnancy: Facts and Controversies. Placenta 2006, 27, 939–958.
- Lyall, F.; Robson, S.C.; Bulmer, J.N. Spiral Artery Remodeling and Trophoblast Invasion in Preeclampsia and Fetal Growth Restriction. Hypertension 2013, 62, 1046–1054.
- Hamilton, W.J.; Boyd, J.D. Trophoblast in Human Utero-Placental Arteries. Nature 1966, 212, 906–908.
- Brosens, I.; Pijnenborg, R.; Vercruysse, L.; Romero, R. The “Great Obstetrical Syndromes” are associated with disorders of deep placentation. Am. J. Obstet. Gynecol. 2011, 204, 193–201.
- Burton, G.J.; Jauniaux, E. Placental Oxidative Stress: From Miscarriage to Preeclampsia. J. Soc. Gynecol. Investig. 2004, 11, 342–352.
- Ball, E.; Bulmer, J.; Ayis, S.; Lyall, F.; Robson, S. Late sporadic miscarriage is associated with abnormalities in spiral artery transformation and trophoblast invasion. J. Pathol. 2006, 208, 535–542.
- Kim, Y.M.; Bujold, E.; Chaiworapongsa, T.; Gomez, R.; Yoon, B.H.; Thaler, H.T.; Rotmensch, S.; Romero, R. Failure of Physiologic Transformation of the Spiral Arteries in Patients with Preterm Labor and Intact Membranes. Am. J. Obstet. Gynecol. 2003, 189, 1063–1069.
- Khong, T.Y.; De Wolf, F.; Robertson, W.B.; Brosens, I. Inadequate Maternal Vascular Response to Placentation in Pregnancies Complicated by Pre-Eclampsia and by Small-for-Gestational Age Infants. Br. J. Obstet. Gynaecol. 1986, 93, 1049–1059.
- Veerbeek, J.H.W.; Nikkels, P.G.J.; Torrance, H.L.; Gravesteijn, J.; Post Uiterweer, E.D.; Derks, J.B.; Koenen, S.V.; Visser, G.H.A.; Van Rijn, B.B.; Franx, A. Placental Pathology in Early Intrauterine Growth Restriction Associated with Maternal Hypertension. Placenta 2014, 35, 696–701.
- Pathak, S.; Lees, C.C.; Hackett, G.; Jessop, F.; Sebire, N.J. Frequency and Clinical Significance of Placental Histological Lesions in an Unselected Population at or near Term. Virchows Arch. 2011, 459, 565–572.
- Maynard, S.E.; Min, J.-Y.; Merchan, J.; Lim, K.-H.; Li, J.; Mondal, S.; Libermann, T.A.; Morgan, J.P.; Sellke, F.W.; Stillman, I.E.; et al. Excess Placental Soluble Fms-like Tyrosine Kinase 1 (sFlt1) May Contribute to Endothelial Dysfunction, Hypertension, and Proteinuria in Preeclampsia. J. Clin. Investig. 2003, 111, 649–658.
- Levine, R.J.; Lam, C.; Qian, C.; Yu, K.F.; Maynard, S.E.; Sachs, B.P.; Sibai, B.M.; Epstein, F.H.; Romero, R.; Thadhani, R.; et al. Soluble Endoglin and Other Circulating Antiangiogenic Factors in Preeclampsia. N. Engl. J. Med. 2006, 355, 992–1005.
- Zeisler, H.; Llurba, E.; Chantraine, F.; Vatish, M.; Staff, A.C.; Sennström, M.; Olovsson, M.; Brennecke, S.P.; Stepan, H.; Allegranza, D.; et al. Predictive Value of the sFlt-1:PlGF Ratio in Women with Suspected Preeclampsia. N. Engl. J. Med. 2016, 374, 13–22.
- Ives, C.W.; Sinkey, R.; Rajapreyar, I.; Tita, A.T.N.; Oparil, S. Preeclampsia—Pathophysiology and Clinical Presentations: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2020, 76, 1690–1702.
- Huppertz, B.; Sammar, M.; Chefetz, I.; Neumaier-Wagner, P.; Bartz, C.; Meiri, H. Longitudinal Determination of Serum Placental Protein 13 during Development of Preeclampsia. Fetal Diagn. Ther. 2008, 24, 230–236.
- Craven, C.M.; Morgan, T.; Ward, K. Decidual Spiral Artery Remodelling Begins before Cellular Interaction with Cytotrophoblasts. Placenta 1998, 19, 241–252.
- Vikse, B.E.; Irgens, L.M.; Leivestad, T.; Skjaerven, R.; Iversen, B.M. Preeclampsia and the Risk of End-Stage Renal Disease. N. Engl. J. Med. 2008, 359, 800–809.
- Kattah, A. Preeclampsia and Kidney Disease: Deciphering Cause and Effect. Curr. Hypertens. Rep. 2020, 22, 91.
- Wiles, K.; Chappell, L.C.; Lightstone, L.; Bramham, K. Updates in Diagnosis and Management of Preeclampsia in Women with CKD. Clin. J. Am. Soc. Nephrol. 2020, 15, 1371–1380.
- Hirose, N.; Ohkuchi, A.; Matsubara, S.; Suzuki, M. Risk of Preeclampsia in Women with CKD, Dialysis or Kidney Transplantation. Med. J. Obstet. Gynecol. 2014, 2, 1028.
- Kendrick, J.; Holmen, J.; You, Z.; Smits, G.; Chonchol, M. Association of Unilateral Renal Agenesis with Adverse Outcomes in Pregnancy: A Matched Cohort Study. Am. J. Kidney Dis. 2017, 70, 506–511.
- Dupont, V.; Berg, A.H.; Yamashita, M.; Huang, C.; Covarrubias, A.E.; Ali, S.; Stotland, A.; Van Eyk, J.E.; Jim, B.; Thadhani, R.; et al. Impaired Renal Reserve Contributes to Preeclampsia via the Kynurenine and Soluble Fms–like Tyrosine Kinase 1 Pathway. J. Clin. Investig. 2022, 132, e158346.
- Chiti, F.; Dobson, C.M. Protein Misfolding, Amyloid Formation, and Human Disease: A Summary of Progress over the Last Decade. Annu. Rev. Biochem. 2017, 86, 27–68.
- Burton, G.J.; Yung, H.-W.; Cindrova-Davies, T.; Charnock-Jones, D.S. Placental Endoplasmic Reticulum Stress and Oxidative Stress in the Pathophysiology of Unexplained Intrauterine Growth Restriction and Early Onset Preeclampsia. Placenta 2009, 30, 43–48.
- Saibil, H. Chaperone Machines for Protein Folding, Unfolding and Disaggregation. Nat. Rev. Mol. Cell Biol. 2013, 14, 630–642.
- Tran, M.; Reddy, P.H. Defective Autophagy and Mitophagy in Aging and Alzheimer’s Disease. Front. Neurosci. 2021, 14, 612757.
- Cuervo, A.M.; Bergamini, E.; Brunk, U.T.; Dröge, W.; Ffrench, M.; Terman, A. Autophagy and Aging: The Importance of Maintaining “Clean” Cells. Autophagy 2005, 1, 131–140.
- Anfinsen, C.B. Principles That Govern the Folding of Protein Chains. Science 1973, 181, 223–230.
- Dobson, C.M. The Structural Basis of Protein Folding and Its Links with Human Disease. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 133–145.
- Dobson, C.M. Protein Folding and Misfolding. Nature 2003, 426, 884–890.
- Knowles, T.P.J.; Vendruscolo, M.; Dobson, C.M. The Amyloid State and Its Association with Protein Misfolding Diseases. Nat. Rev. Mol. Cell Biol. 2014, 15, 384–396.
- Newberry, R.W.; Raines, R.T. Secondary Forces in Protein Folding. ACS Chem. Biol. 2019, 14, 1677.
- Dill, K.A. Dominant Forces in Protein Folding. Biochemistry 1990, 29, 7133–7155.
- Kang, T.S.; Kini, R.M. Structural Determinants of Protein Folding. Cell. Mol. Life Sci. 2009, 66, 2341–2361.
- Dobson, C.M. Principles of Protein Folding, Misfolding and Aggregation. Semin. Cell Dev. Biol. 2004, 15, 3–16.
- Vendruscolo, M.; Knowles, T.P.J.; Dobson, C.M. Protein Solubility and Protein Homeostasis: A Generic View of Protein Misfolding Disorders. Cold Spring Harb. Perspect. Biol. 2011, 3, a010454.
- Kelly, J.W. The Alternative Conformations of Amyloidogenic Proteins and Their Multi-Step Assembly Pathways. Curr. Opin. Struct. Biol. 1998, 8, 101–106.
- Cohen, S.I.A.; Linse, S.; Luheshi, L.M.; Hellstrand, E.; White, D.A.; Rajah, L.; Otzen, D.E.; Vendruscolo, M.; Dobson, C.M.; Knowles, T.P.J. Proliferation of Amyloid-Β42 Aggregates Occurs through a Secondary Nucleation Mechanism. Proc. Natl. Acad. Sci. USA 2013, 110, 9758–9763.
- Knowles, T.P.J.; White, D.A.; Abate, A.R.; Agresti, J.J.; Cohen, S.I.A.; Sperling, R.A.; De Genst, E.J.; Dobson, C.M.; Weitz, D.A. Observation of Spatial Propagation of Amyloid Assembly from Single Nuclei. Proc. Natl. Acad. Sci. USA 2011, 108, 14746–14751.
- Cohen, S.I.A.; Vendruscolo, M.; Dobson, C.M.; Knowles, T.P.J. From Macroscopic Measurements to Microscopic Mechanisms of Protein Aggregation. J. Mol. Biol. 2012, 421, 160–171.
- Knowles, T.P.J.; Waudby, C.A.; Devlin, G.L.; Cohen, S.I.A.; Aguzzi, A.; Vendruscolo, M.; Terentjev, E.M.; Welland, M.E.; Dobson, C.M. An Analytical Solution to the Kinetics of Breakable Filament Assembly. Science 2009, 326, 1533–1537.
- Cline, E.N.; Bicca, M.A.; Viola, K.L.; Klein, W.L. The Amyloid-β Oligomer Hypothesis: Beginning of the Third Decade. J. Alzheimers Dis. 2018, 64, S567–S610.
- Koike, H.; Iguchi, Y.; Sahashi, K.; Katsuno, M. Significance of Oligomeric and Fibrillar Species in Amyloidosis: Insights into Pathophysiology and Treatment. Molecules 2021, 26, 5091.
- Buhimschi, I.A.; Zhao, G.; Funai, E.F.; Harris, N.; Sasson, I.E.; Bernstein, I.M.; Saade, G.R.; Buhimschi, C.S. Proteomic profiling of urine identifies specific fragments of Serpina-1 and albumin as biomarkers of preeclampsia. Am. J. Obstet. Gynecol. 2008, 199, 551.e1–551.e16.
- Buhimschi, I.A.; Nayeri, U.A.; Zhao, G.; Shook, L.L.; Pensalfini, A.; Funai, E.F.; Bernstein, I.M.; Glabe, C.G.; Buhimschi, C.S. Protein Misfolding, Congophilia, Oligomerization, and Defective Amyloid Processing in Preeclampsia. Sci. Transl. Med. 2014, 6, 245ra92.
- Puchtler, H.; Sweat, F.; Levine, M. On the binding of congo red by amyloid. J. Histochem. Cytochem. 1962, 10, 355–364.
- Kalkunte, S.S.; Neubeck, S.; Norris, W.E.; Cheng, S.-B.; Kostadinov, S.; Vu Hoang, D.; Ahmed, A.; von Eggeling, F.; Shaikh, Z.; Padbury, J.; et al. Transthyretin Is Dysregulated in Preeclampsia, and Its Native Form Prevents the Onset of Disease in a Preclinical Mouse Model. Am. J. Pathol. 2013, 183, 1425–1436.
- Cheng, S.; Banerjee, S.; Daiello, L.A.; Nakashima, A.; Jash, S.; Huang, Z.; Drake, J.D.; Ernerudh, J.; Berg, G.; Padbury, J.; et al. Novel Blood Test for Early Biomarkers of Preeclampsia and Alzheimer’s Disease. Sci. Rep. 2021, 11, 15934.
- Carrell, R.W.; Mushunje, A.; Zhou, A. Serpins Show Structural Basis for Oligomer Toxicity and Amyloid Ubiquity. FEBS Lett. 2008, 582, 2537–2541.
- Lewandowski, C.T.; Weng, J.M.; LaDu, M.J. Alzheimer’s Disease Pathology in APOE Transgenic Mouse Models: The Who, What, When, Where, Why, and How. Neurobiol. Dis. 2020, 139, 104811.
- Medeiros, R.; Baglietto-Vargas, D.; LaFerla, F.M. The Role of Tau in Alzheimer’s Disease and Related Disorders. CNS Neurosci. Ther. 2010, 17, 514–524.
- Tong, M.; Cheng, S.; Chen, Q.; DeSousa, J.; Stone, P.R.; James, J.L.; Chamley, L.W.; Sharma, S. Aggregated Transthyretin Is Specifically Packaged into Placental Nano-Vesicles in Preeclampsia. Sci. Rep. 2017, 7, 6694.
- Roussel, B.D.; Irving, J.A.; Ekeowa, U.I.; Belorgey, D.; Haq, I.; Ordóñez, A.; Kruppa, A.J.; Duvoix, A.; Rashid, S.T.; Crowther, D.C.; et al. Unravelling the Twists and Turns of the Serpinopathies. FEBS J. 2011, 278, 3859–3867.
- Köhnlein, T.; Welte, T. Alpha-1 Antitrypsin Deficiency: Pathogenesis, Clinical Presentation, Diagnosis, and Treatment. Am. J. Med. 2008, 121, 3–9.
- Yuan, Y.; DiCiaccio, B.; Li, Y.; Elshikha, A.S.; Titov, D.; Brenner, B.; Seifer, L.; Pan, H.; Karic, N.; Akbar, M.A.; et al. Anti-inflammaging Effects of Human Alpha-1 Antitrypsin. Aging Cell 2018, 17, e12694.
- Starodubtseva, N.; Nizyaeva, N.; Baev, O.; Bugrova, A.; Gapaeva, M.; Muminova, K.; Kononikhin, A.; Frankevich, V.; Nikolaev, E.; Sukhikh, G. SERPINA1 Peptides in Urine as A Potential Marker of Preeclampsia Severity. Int. J. Mol. Sci. 2020, 21, 914.
- Cheng, S.-B.; Nakashima, A.; Huber, W.J.; Davis, S.; Banerjee, S.; Huang, Z.; Saito, S.; Sadovsky, Y.; Sharma, S. Pyroptosis Is a Critical Inflammatory Pathway in the Placenta from Early Onset Preeclampsia and in Human Trophoblasts Exposed to Hypoxia and Endoplasmic Reticulum Stressors. Cell Death Dis. 2019, 10, 927.
- Twina, G.; Sheiner, E.; Shahaf, G.; Yaniv Salem, S.; Madar, T.; Baron, J.; Wiznitzer, A.; Mazor, M.; Holcberg, G.; Lewis, E.C. Lower Circulation Levels and Activity of α-1 Antitrypsin in Pregnant Women with Severe Preeclampsia. J. Matern.-Fetal Neonatal Med. 2012, 25, 2667–2670.
- Feng, Y.; Xu, J.; Zhou, Q.; Wang, R.; Liu, N.; Wu, Y.; Yuan, H.; Che, H. Alpha-1 Antitrypsin Prevents the Development of Preeclampsia Through Suppression of Oxidative Stress. Front. Physiol. 2016, 7, 176.
- Feng, Y.; Wang, N.; Xu, J.; Zou, J.; Liang, X.; Liu, H.; Chen, Y. Alpha-1-Antitrypsin Functions as a Protective Factor in Preeclampsia through Activating Smad2 and Inhibitor of DNA Binding 4. Oncotarget 2017, 8, 113002–113012.
- Nagarajappa, C.; Rangappa, S.S.; Balakrishna, S. Misfolding Linked Mutations of SERPINA1 Gene Are Uncommon in Preeclampsia. Arch. Med. Health Sci. 2019, 7, 177.
- Shaik, N.A.; Saud Al-Saud, N.B.; Abdulhamid Aljuhani, T.; Jamil, K.; Alnuman, H.; Aljeaid, D.; Sultana, N.; El-Harouni, A.A.; Awan, Z.A.; Elango, R.; et al. Structural Characterization and Conformational Dynamics of Alpha-1 Antitrypsin Pathogenic Variants Causing Alpha-1-Antitrypsin Deficiency. Front. Mol. Biosci. 2022, 9, 1051511.
- Sergeeva, V.A.; Zakharova, N.V.; Bugrova, A.E.; Starodubtseva, N.L.; Indeykina, M.I.; Kononikhin, A.S.; Frankevich, V.E.; Nikolaev, E.N. The High-Resolution Mass Spectrometry Study of the Protein Composition of Amyloid-like Urine Aggregates Associated with Preeclampsia. Eur. J. Mass. Spectrom. 2020, 26, 158–161.
- Valtanen, R.S.; Buhimschi, C.S.; Bahtiyar, M.O.; Zhao, G.; Jing, H.; Ackerman, W.E.; Glabe, C.G.; Buhimschi, I.A. Conformation-Dependent Anti-Aβ Monoclonal Antibody Signatures of Disease Status and Severity in Urine of Women with Preeclampsia. Pregnancy Hypertens. 2022, 28, 51–59.
- Sun, X.; Chen, W.-D.; Wang, Y.-D. β-Amyloid: The Key Peptide in the Pathogenesis of Alzheimer’s Disease. Front. Pharmacol. 2015, 6, 221.
- Müller, U.C.; Deller, T.; Korte, M. Not Just Amyloid: Physiological Functions of the Amyloid Precursor Protein Family. Nat. Rev. Neurosci. 2017, 18, 281–298.
- Chen, G.; Xu, T.; Yan, Y.; Zhou, Y.; Jiang, Y.; Melcher, K.; Xu, H.E. Amyloid Beta: Structure, Biology and Structure-Based Therapeutic Development. Acta Pharmacol. Sin. 2017, 38, 1205–1235.
- Xia, Y.; Prokop, S.; Giasson, B.I. “Don’t Phos Over Tau”: Recent Developments in Clinical Biomarkers and Therapies Targeting Tau Phosphorylation in Alzheimer’s Disease and Other Tauopathies. Mol. Neurodegener. 2021, 16, 37.
- Guo, T.; Noble, W.; Hanger, D.P. Roles of Tau Protein in Health and Disease. Acta Neuropathol. 2017, 133, 665–704.
- Liz, M.A.; Coelho, T.; Bellotti, V.; Fernandez-Arias, M.I.; Mallaina, P.; Obici, L. A Narrative Review of the Role of Transthyretin in Health and Disease. Neurol. Ther. 2020, 9, 395–402.
- Patel, J.; Landers, K.A.; Li, H.; Mortimer, R.H.; Richard, K. Ontogenic Changes in Placental Transthyretin. Placenta 2011, 32, 817–822.
- Gião, T.; Saavedra, J.; Cotrina, E.; Quintana, J.; Llop, J.; Arsequell, G.; Cardoso, I. Undiscovered Roles for Transthyretin: From a Transporter Protein to a New Therapeutic Target for Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 2075.
- Nunes, R.J.; de Oliveira, P.; Lages, A.; Becker, J.D.; Marcelino, P.; Barroso, E.; Perdigoto, R.; Kelly, J.W.; Quintas, A.; Santos, S.C.R. Transthyretin Proteins Regulate Angiogenesis by Conferring Different Molecular Identities to Endothelial Cells*. J. Biol. Chem. 2013, 288, 31752–31760.
- Reixach, N.; Deechongkit, S.; Jiang, X.; Kelly, J.W.; Buxbaum, J.N. Tissue Damage in the Amyloidoses: Transthyretin Monomers and Nonnative Oligomers Are the Major Cytotoxic Species in Tissue Culture. Proc. Natl. Acad. Sci. USA 2004, 101, 2817–2822.
- Burton, G.J.; Yung, H.-W. Endoplasmic Reticulum Stress in the Pathogenesis of Early-Onset Pre-Eclampsia. Pregnancy Hypertens. 2011, 1, 72–78.
- Hemagirri, M.; Chen, Y.; Gopinath, S.C.B.; Sahreen, S.; Adnan, M.; Sasidharan, S. Crosstalk between Protein Misfolding and Endoplasmic Reticulum Stress during Ageing and Their Role in Age-Related Disorders. Biochimie 2023.
- Yoshida, H. ER Stress and Diseases. FEBS J. 2007, 274, 630–658.
- Cheng, S.; Huang, Z.; Banerjee, S.; Jash, S.; Buxbaum, J.N.; Sharma, S. Evidence from Human Placenta, Endoplasmic Reticulum–Stressed Trophoblasts, and Transgenic Mice Links Transthyretin Proteinopathy to Preeclampsia. Hypertension 2022, 79, 1738–1754.
- Du, L.; He, F.; Kuang, L.; Tang, W.; Li, Y.; Chen, D. eNOS/iNOS and Endoplasmic Reticulum Stress-Induced Apoptosis in the Placentas of Patients with Preeclampsia. J. Hum. Hypertens. 2017, 31, 49–55.
- Expression of Markers of Endoplasmic Reticulum Stress-Induced Apoptosis in the Placenta of Women with Early and Late Onset Severe Pre-Eclampsia—ClinicalKey. Available online: https://www.clinicalkey.com/#!/content/playContent/1-s2.0-S1028455914002022?returnurl=https:%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS1028455914002022%3Fshowall%3Dtrue&referrer=https:%2F%2Fpubmed.ncbi.nlm.nih.gov%2F (accessed on 22 November 2023).
- Lian, I.A.; Løset, M.; Mundal, S.B.; Fenstad, M.H.; Johnson, M.P.; Eide, I.P.; Bjørge, L.; Freed, K.A.; Moses, E.K.; Austgulen, R. Increased Endoplasmic Reticulum Stress in Decidual Tissue from Pregnancies Complicated by Fetal Growth Restriction with and without Pre-Eclampsia. Placenta 2011, 32, 823–829.
- Yung, H.; Calabrese, S.; Hynx, D.; Hemmings, B.A.; Cetin, I.; Charnock-Jones, D.S.; Burton, G.J. Evidence of Placental Translation Inhibition and Endoplasmic Reticulum Stress in the Etiology of Human Intrauterine Growth Restriction. Am. J. Pathol. 2008, 173, 451–462.
- Mochan, S.; Dhingra, M.K.; Gupta, S.K.; Saxena, S.; Arora, P.; Yadav, V.; Rani, N.; Luthra, K.; Dwivedi, S.; Bhatla, N.; et al. Status of VEGF in Preeclampsia and Its Effect on Endoplasmic Reticulum Stress in Placental Trophoblast Cells. Eur. J. Obstet. Gynecol. Reprod. Biol. X 2019, 4, 100070.
- Iyer, K.; Chand, K.; Mitra, A.; Trivedi, J.; Mitra, D. Diversity in Heat Shock Protein Families: Functional Implications in Virus Infection with a Comprehensive Insight of Their Role in the HIV-1 Life Cycle. Cell Stress. Chaperones 2021, 26, 743–768.
- Tripathi, A.; Iyer, K.; Mitra, D. HIV-1 Replication Requires Optimal Activation of the Unfolded Protein Response. FEBS Lett. 2023, 597, 2908–2930.
- Jee, B.; Dhar, R.; Singh, S.; Karmakar, S. Heat Shock Proteins and Their Role in Pregnancy: Redefining the Function of “Old Rum in a New Bottle”. Front. Cell Dev. Biol. 2021, 9, 648463.
- Rai, S.; Tapadia, M.G. Hsc70-4 Aggravates PolyQ-Mediated Neurodegeneration by Modulating NF-κB Mediated Immune Response in Drosophila. Front. Mol. Neurosci. 2022, 15, 857257.
- Cater, J.H.; Kumita, J.R.; Zeineddine Abdallah, R.; Zhao, G.; Bernardo-Gancedo, A.; Henry, A.; Winata, W.; Chi, M.; Grenyer, B.S.F.; Townsend, M.L.; et al. Human Pregnancy Zone Protein Stabilizes Misfolded Proteins Including Preeclampsia- and Alzheimer’s-Associated Amyloid Beta Peptide. Proc. Natl. Acad. Sci. USA 2019, 116, 6101–6110.
- Devriendt, K.; Van den Berghe, H.; Cassiman, J.-J.; Marynen, P. Primary Structure of Pregnancy Zone Protein. Molecular Cloning of a Full-Length PZP cDNA Clone by the Polymerase Chain Reaction. Biochim. Biophys. Acta (BBA)-Gene Struct. Expr. 1991, 1088, 95–103.
- Sottrup-Jensen, L.; Folkersen, J.; Kristensen, T.; Tack, B.F. Partial Primary Structure of Human Pregnancy Zone Protein: Extensive Sequence Homology with Human Alpha 2-Macroglobulin. Proc. Natl. Acad. Sci. USA 1984, 81, 7353–7357.
- Folkersen, J.; Teisner, B.; Grunnet, N.; Grudzinskas, J.G.; Westergaard, J.G.; Hindersson, P. Circulating Levels of Pregnancy Zone Protein: Normal Range and the Influence of Age and Gender. Clin. Chim. Acta 1981, 110, 139–145.
- Sand, O.; Folkersen, J.; Westergaard, J.G.; Sottrup-Jensen, L. Characterization of Human Pregnancy Zone Protein. Comparison with Human Alpha 2-Macroglobulin. J. Biol. Chem. 1985, 260, 15723–15735.
- Cater, J.H.; Mañucat-Tan, N.B.; Georgiou, D.K.; Zhao, G.; Buhimschi, I.A.; Wyatt, A.R.; Ranson, M. A Novel Role for Plasminogen Activator Inhibitor Type-2 as a Hypochlorite-Resistant Serine Protease Inhibitor and Holdase Chaperone. Cells 2022, 11, 1152.
- Wyatt, A.R.; Cater, J.H.; Ranson, M. PZP and PAI-2: Structurally-Diverse, Functionally Similar Pregnancy Proteins? Int. J. Biochem. Cell Biol. 2016, 79, 113–117.
- Ohkuchi, A.; Minakami, H.; Aoya, T.; Haga, T.; Kimura, H.; Suzuki, M.; Sato, I. Expansion of the Fraction of Th1 Cells in Women with Preeclampsia: Inverse Correlation between the Percentage of Th1 Cells and the Plasma Level of PAI-2. Am. J. Reprod. Immunol. 2001, 46, 252–259.
- Saito, S.; Shima, T.; Nakashima, A. Immunological Maladaptation. In Preeclampsia: Basic, Genomic, and Clinical; Saito, S., Ed.; Comprehensive Gynecology and Obstetrics; Springer: Singapore, 2018; pp. 65–84. ISBN 978-981-10-5891-2.
- Grancha, S.; Estellés, A.; Gilabert, J.; Chirivella, M.; España, F.; Aznar, J. Decreased Expression of PAI-2 mRNA and Protein in Pregnancies Complicated with Intrauterine Fetal Growth Retardation. Thromb. Haemost. 1996, 76, 761–767.
- Roes, E.M.; Sweep, C.G.F.; Thomas, C.M.; Zusterzeel, P.L.; Geurts-Moespot, A.; Peters, W.H.; Steegers, E.A. Levels of Plasminogen Activators and Their Inhibitors in Maternal and Umbilical Cord Plasma in Severe Preeclampsia. Am. J. Obstet. Gynecol. 2002, 187, 1019–1025.
- de Graff, A.M.; Mosedale, D.E.; Sharp, T.; Dill, K.A.; Grainger, D.J. Proteostasis Is Adaptive: Balancing Chaperone Holdases against Foldases. PLoS Comput. Biol. 2020, 16, e1008460.
- Lee, J.A.; Yerbury, J.J.; Farrawell, N.; Shearer, R.F.; Constantinescu, P.; Hatters, D.M.; Schroder, W.A.; Suhrbier, A.; Wilson, M.R.; Saunders, D.N.; et al. SerpinB2 (PAI-2) Modulates Proteostasis via Binding Misfolded Proteins and Promotion of Cytoprotective Inclusion Formation. PLoS ONE 2015, 10, e0130136.
- Nakashima, A.; Cheng, S.-B.; Ikawa, M.; Yoshimori, T.; Huber, W.J.; Menon, R.; Huang, Z.; Fierce, J.; Padbury, J.F.; Sadovsky, Y.; et al. Evidence for Lysosomal Biogenesis Proteome Defect and Impaired Autophagy in Preeclampsia. Autophagy 2020, 16, 1771–1785.
- Cheng, S.-B.; Nakashima, A.; Sharma, S. Understanding Pre-Eclampsia Using Alzheimer’s Etiology: An Intriguing Viewpoint. Am. J. Reprod. Immunol. 2016, 75, 372–381.
- Nakashima, A.; Cheng, S.-B.; Kusabiraki, T.; Motomura, K.; Aoki, A.; Ushijima, A.; Ono, Y.; Tsuda, S.; Shima, T.; Yoshino, O.; et al. Endoplasmic Reticulum Stress Disrupts Lysosomal Homeostasis and Induces Blockade of Autophagic Flux in Human Trophoblasts. Sci. Rep. 2019, 9, 11466.
- Nakashima, A.; Tsuda, S.; Kusabiraki, T.; Aoki, A.; Ushijima, A.; Shima, T.; Cheng, S.-B.; Sharma, S.; Saito, S. Current Understanding of Autophagy in Pregnancy. Int. J. Mol. Sci. 2019, 20, 2342.
- Nakashima, A.; Shima, T.; Tsuda, S.; Aoki, A.; Kawaguchi, M.; Furuta, A.; Yasuda, I.; Yoneda, S.; Yamaki-Ushijima, A.; Cheng, S.-B.; et al. Aggrephagy Deficiency in the Placenta: A New Pathogenesis of Preeclampsia. Int. J. Mol. Sci. 2021, 22, 2432.
- Nakashima, A.; Yamanaka-Tatematsu, M.; Fujita, N.; Koizumi, K.; Shima, T.; Yoshida, T.; Nikaido, T.; Okamoto, A.; Yoshimori, T.; Saito, S. Impaired Autophagy by Soluble Endoglin, under Physiological Hypoxia in Early Pregnant Period, Is Involved in Poor Placentation in Preeclampsia. Autophagy 2013, 9, 303–316.
- Parzych, K.R.; Klionsky, D.J. An Overview of Autophagy: Morphology, Mechanism, and Regulation. Antioxid. Redox Signal 2014, 20, 460–473.
- Opoku-Nsiah, K.A.; Gestwicki, J.E. Aim for the Core: Suitability of the Ubiquitin-Independent 20S Proteasome as a Drug Target in Neurodegeneration. Transl. Res. 2018, 198, 48–57.
- Nakashima, A.; Shima, T.; Aoki, A.; Kawaguchi, M.; Yasuda, I.; Tsuda, S.; Yoneda, S.; Yamaki-Ushijima, A.; Cheng, S.; Sharma, S.; et al. Placental Autophagy Failure: A Risk Factor for Preeclampsia. J. Obstet. Gynaecol. Res. 2020, 46, 2497–2504.
- Ye, H.; Chen, M.; Cao, F.; Huang, H.; Zhan, R.; Zheng, X. Chloroquine, an Autophagy Inhibitor, Potentiates the Radiosensitivity of Glioma Initiating Cells by Inhibiting Autophagy and Activating Apoptosis. BMC Neurol. 2016, 16, 178.
- Owaraganise, A.; Migisha, R.; Ssalongo, W.G.M.; Tibaijuka, L.; Kayondo, M.; Twesigomwe, G.; Ngonzi, J.; Lugobe, H.M. Nonproteinuric Preeclampsia among Women with Hypertensive Disorders of Pregnancy at a Referral Hospital in Southwestern Uganda. Obstet. Gynecol. Int. 2021, 2021, 9751775.
- Palmqvist, S.; Insel, P.S.; Stomrud, E.; Janelidze, S.; Zetterberg, H.; Brix, B.; Eichenlaub, U.; Dage, J.L.; Chai, X.; Blennow, K.; et al. Cerebrospinal Fluid and Plasma Biomarker Trajectories with Increasing Amyloid Deposition in Alzheimer’s Disease. EMBO Mol. Med. 2019, 11, e11170.
- Schindler, S.E.; Bollinger, J.G.; Ovod, V.; Mawuenyega, K.G.; Li, Y.; Gordon, B.A.; Holtzman, D.M.; Morris, J.C.; Benzinger, T.L.S.; Xiong, C.; et al. High-Precision Plasma β-Amyloid 42/40 Predicts Current and Future Brain Amyloidosis. Neurology 2019, 93, e1647–e1659.
- Nakamura, A.; Kaneko, N.; Villemagne, V.L.; Kato, T.; Doecke, J.; Doré, V.; Fowler, C.; Li, Q.-X.; Martins, R.; Rowe, C.; et al. High Performance Plasma Amyloid-β Biomarkers for Alzheimer’s Disease. Nature 2018, 554, 249–254.
- Stevenson-Hoare, J.; Heslegrave, A.; Leonenko, G.; Fathalla, D.; Bellou, E.; Luckcuck, L.; Marshall, R.; Sims, R.; Morgan, B.P.; Hardy, J.; et al. Plasma Biomarkers and Genetics in the Diagnosis and Prediction of Alzheimer’s Disease. Brain 2023, 146, 690–699.
- Palmqvist, S.; Tideman, P.; Cullen, N.; Zetterberg, H.; Blennow, K.; Dage, J.L.; Stomrud, E.; Janelidze, S.; Mattsson-Carlgren, N.; Hansson, O. Prediction of Future Alzheimer’s Disease Dementia Using Plasma Phospho-Tau Combined with Other Accessible Measures. Nat. Med. 2021, 27, 1034–1042.
- McCarthy, F.P.; Adetoba, A.; Gill, C.; Bramham, K.; Bertolaccini, M.; Burton, G.J.; Girardi, G.; Seed, P.T.; Poston, L.; Chappell, L.C. Urinary Congophilia in Women with Hypertensive Disorders of Pregnancy and Preexisting Proteinuria or Hypertension. Am. J. Obstet. Gynecol. 2016, 215, 464.e1–464.e7.
- Nagarajappa, C.; Rangappa, S.S.; Suryanarayana, R.; Balakrishna, S. Urinary Congophilia in Preeclampsia: Experience from a Rural Tertiary-Care Hospital in India. Pregnancy Hypertens. 2018, 13, 83–86.
- Khaliq, O.P.; Phoswa, W.N.; Moodley, J. The Effectiveness of the Congo Red Dot Paper Test in Hypertensive Disorders of Pregnancy: A Systematic Review and Meta-Analysis. Front. Reprod. Health 2023, 5, 1120937.
- Wong, N.K.L.; Wah, I.Y.M.; Wong, S.T.K.; Nguyen-Hoang, L.; Lau, C.S.L.; Ip, P.N.P.; Leung, H.H.Y.; Sahota, D.S.; Poon, L.C. A Point-of-Care Urine Test to Predict Adverse Maternal and Neonatal Outcomes in Asian Women with Suspected Preeclampsia. Arch. Gynecol. Obstet. 2023, 1–10.
- Wong, S.T.K.; Sahota, D.S.; Wong, N.K.L.; Wah, I.Y.M.; Wang, X.; Lau, S.L.; Chiu, C.P.H.; Ip, P.N.P.; Poon, L.C. A Point-of Care Urine Test to Predict Preeclampsia Development in Asian Women with Suspected Preeclampsia. Pregnancy Hypertens. 2023, 32, 28–34.
- Sailakshmi, M.P.A.; Prabhu, M.R.; Prabhakara, S.; Anbazhagan, K.; Rupakala, B.M. Congo Red Dot Test in the Early Prediction and Diagnosis of Pre-Eclampsia in a Tertiary Health Care Centre in India. Pregnancy Hypertens. 2021, 25, 225–229.
- Sammar, M.; Syngelaki, A.; Sharabi-Nov, A.; Nicolaides, K.; Meiri, H. Can Staining of Damaged Proteins in Urine Effectively Predict Preeclampsia? Fetal Diagn. Ther. 2017, 41, 23–31.
- Younis, D.; Mosbah, A.; Zakaria, M.M.; Awadalla, A.; El-Kannishy, G.; Shemies, R.S. Urinary Congophilia in Pregnancy: A Marker of Kidney Injury Rather than Preeclampsia. J. Hypertens. 2023, 41, 1760–1767.
- Döbert, M.; Varouxaki, A.-N.; Mu, A.C.; Syngelaki, A.; Nicolaides, K.H. Screening for Late Preeclampsia at 35-37 Weeks by the Urinary Congo-Red Dot Paper Test. J. Matern. Fetal Neonatal Med. 2022, 35, 5686–5690.
- Li, X.-M.; Liu, X.-M.; Xu, J.; Du, J.; Cuckle, H. Late Pregnancy Screening for Preeclampsia with a Urinary Point-of-Care Test for Misfolded Proteins. PLoS ONE 2020, 15, e0233214.
- Bracken, H.; Buhimschi, I.A.; Rahman, A.; Smith, P.R.S.; Pervin, J.; Rouf, S.; Bousieguez, M.; López, L.G.; Buhimschi, C.S.; Easterling, T.; et al. Congo Red Test for Identification of Preeclampsia: Results of a Prospective Diagnostic Case-Control Study in Bangladesh and Mexico. EClinicalMedicine 2021, 31, 100678.
- Rood, K.M.; Buhimschi, C.S.; Dible, T.; Webster, S.; Zhao, G.; Samuels, P.; Buhimschi, I.A. Congo Red Dot Paper Test for Antenatal Triage and Rapid Identification of Preeclampsia. EClinicalMedicine 2019, 8, 47–56.
- Millen, K.R.; Buhimschi, C.S.; Zhao, G.; Rood, K.M.; Tabbah, S.; Buhimschi, I.A. Serum and urine thioflavin T (ThT) enhanced fluorescence in severe preeclampsia. Hypertension 2018, 71, 1185–1192.
- Rodriguez Chavez, J.L.; Fuentes Gutiérrez, E.K.; de Jesús Angeles Vázquez, M.; Mendieta Zerón, H. Evaluation of Congo Red Staining Kit to Determine Proteinuria in Preeclampsia. J. Clin. Diagn. Res. 2018, 12, QM01–QM05.
- Planté-Bordeneuve, V.; Said, G. Familial Amyloid Polyneuropathy. Lancet Neurol. 2011, 10, 1086–1097.
- Cardoso, I.; Goldsbury, C.S.; Müller, S.A.; Olivieri, V.; Wirtz, S.; Damas, A.M.; Aebi, U.; Saraiva, M.J. Transthyretin Fibrillogenesis Entails the Assembly of Monomers: A Molecular Model for in Vitro Assembled Transthyretin Amyloid-like Fibrils. J. Mol. Biol. 2002, 317, 683–695.
- Cotrina, E.Y.; Santos, L.M.; Rivas, J.; Blasi, D.; Leite, J.P.; Liz, M.A.; Busquets, M.A.; Planas, A.; Prohens, R.; Gimeno, A.; et al. Targeting Transthyretin in Alzheimer’s Disease: Drug Discovery of Small-Molecule Chaperones as Disease-Modifying Drug Candidates for Alzheimer’s Disease. Eur. J. Med. Chem. 2021, 226, 113847.
- Ajoolabady, A.; Lindholm, D.; Ren, J.; Pratico, D. ER Stress and UPR in Alzheimer’s Disease: Mechanisms, Pathogenesis, Treatments. Cell Death Dis. 2022, 13, 706.
- Huang, L.-K.; Kuan, Y.-C.; Lin, H.-W.; Hu, C.-J. Clinical Trials of New Drugs for Alzheimer Disease: A 2020–2023 Update. J. Biomed. Sci. 2023, 30, 83.
- Ji, C.; Sigurdsson, E.M. Current Status of Clinical Trials on Tau Immunotherapies. Drugs 2021, 81, 1135–1152.
- Eshraghi, M.; Ahmadi, M.; Afshar, S.; Lorzadeh, S.; Adlimoghaddam, A.; Rezvani Jalal, N.; West, R.; Dastghaib, S.; Igder, S.; Torshizi, S.R.N.; et al. Enhancing Autophagy in Alzheimer’s Disease through Drug Repositioning. Pharmacol. Ther. 2022, 237, 108171.
- Fang, J.; Zhang, P.; Zhou, Y.; Chiang, C.-W.; Tan, J.; Hou, Y.; Stauffer, S.; Li, L.; Pieper, A.A.; Cummings, J.; et al. Endophenotype-Based in Silico Network Medicine Discovery Combined with Insurance Record Data Mining Identifies Sildenafil as a Candidate Drug for Alzheimer’s Disease. Nat. Aging 2021, 1, 1175–1188.
- El-Bakly, W.; Wagdy, O.; Sobhy, A.; Abo Elenain, O.; Riad, M.S.; El Sayed, M.; Tarkhan, S.; Yassen, M.; Mahmoud, A.; Bassiony, M.; et al. The Efficacy and Underlying Mechanism of Phosphodiesterase- 5 Inhibitors in Preventing Cognitive Impairment and Alzheimer Pathology: A Systematic Review of Animal Studies. Behav. Brain Res. 2019, 372, 112004.
- Desai, R.J.; Mahesri, M.; Lee, S.B.; Varma, V.R.; Loeffler, T.; Schilcher, I.; Gerhard, T.; Segal, J.B.; Ritchey, M.E.; Horton, D.B.; et al. No Association between Initiation of Phosphodiesterase-5 Inhibitors and Risk of Incident Alzheimer’s Disease and Related Dementia: Results from the Drug Repurposing for Effective Alzheimer’s Medicines Study. Brain Commun. 2022, 4, fcac247.
- Andraweera, P.H.; Lassi, Z.S. Cardiovascular Risk Factors in Offspring of Preeclamptic Pregnancies-Systematic Review and Meta-Analysis. J. Pediatr. 2019, 208, 104–113.e6.
- Nahum Sacks, K.; Friger, M.; Shoham-Vardi, I.; Spiegel, E.; Sergienko, R.; Landau, D.; Sheiner, E. Prenatal Exposure to Preeclampsia as an Independent Risk Factor for Long-Term Cardiovascular Morbidity of the Offspring. Pregnancy Hypertens. 2018, 13, 181–186.
- Ray, J.G.; Vermeulen, M.J.; Schull, M.J.; Redelmeier, D.A. Cardiovascular Health after Maternal Placental Syndromes (CHAMPS): Population-Based Retrospective Cohort Study. Lancet 2005, 366, 1797–1803.
- Wu, P.; Haththotuwa, R.; Kwok, C.S.; Babu, A.; Kotronias, R.A.; Rushton, C.; Zaman, A.; Fryer, A.A.; Kadam, U.; Chew-Graham, C.A.; et al. Preeclampsia and Future Cardiovascular Health. Circ. Cardiovasc. Qual. Outcomes 2017, 10, e003497.
- Andolf, E.; Bladh, M.; Möller, L.; Sydsjö, G. Prior Placental Bed Disorders and Later Dementia: A Retrospective Swedish Register-Based Cohort Study. BJOG Int. J. Obstet. Gynaecol. 2020, 127, 1090–1099.
- Basit, S.; Wohlfahrt, J.; Boyd, H.A. Pre-Eclampsia and Risk of Dementia Later in Life: Nationwide Cohort Study. BMJ 2018, 363, k4109.