Oral squamous cell carcinoma (OSCC) remains an understudied and significant global cancer killer and dismal survival rates have not changed in decades. A better understanding of the molecular basis of OSCC progression and metastasis is needed to develop new approaches for treating this disease. The supportive network surrounding cancer tumor cells known as the tumor microenvironment (TME) has gained increasing interest lately since it performs essential protumorigenic functions. Cancer-associated fibroblasts (CAFs) are one of the main cell types in the TME and are known to play a key role in influencing the biological behavior of tumors.
- cancer-associated fibroblast
- CAF subtype
- oral squamous cell carcinoma
- tumor microenvironment
1. Introduction
2. Defining Fibroblasts
Fibroblasts exist in almost all tissue types and are the main cell type in stroma that produces the structural framework of human tissue through creation and maintenance of the extracellular matrix (ECM). Fibroblasts are derived mainly from primitive mesenchymal cells, although alternative origins have been reported (this is described in a subsequent section). There is no single marker used to identify fibroblast cells. In practice, fibroblasts taken from tumors/ tissues are usually first defined through negative selection: they do not express epithelial (EpCAM), endothelial (CD31), leukocyte (CD45), or erythrocyte (CD235a) markers. Next, positive expression of mesenchymal markers such as Vimentin (VIM), alpha-smooth muscle actin (αSMA), and others is used to indicate that a given cell is a fibroblast. In culture, fibroblasts can also be characterized by their spindle-like appearance. Fibroblast functions include tissue homeostasis, wound healing response through secretion of extracellular matrix proteins to help heal the area, and/or through secretion of cytokines to recruit immune cells and initiate an inflammatory response. Normal fibroblasts can also serve as progenitors for specialized mesenchymal cell types, such as bone-forming osteoblasts and lipid-filled adipocytes [6][19]. Further, fibroblasts activated in a TME are called CAFs and these are known to display a vast array of functional abilities affecting tumor progression.3. Fibroblast Activation and CAF Markers
3.1. Fibroblast Plasticity
CAFs may originate from normal tissue fibroblasts, as reported by others [7][20], with lineage tracing studies in genetically engineered mouse models supporting this concept [8][21]. These lineage tracing mouse models utilize a specific fibroblast gene promoter attached to a reporter molecule (e.g., mCherry or GFP). This reporter is expressed once the cell begins to express the fibroblast gene used to promote the reporter in the mouse (e.g., a Col5a1 promoter). The cell continues to express the reporter once activated, and expression of the marker is passed down to all daughter cells. This approach has allowed tracking of a chosen cell type as it morphs into a CAF in a tumor setting. An example of a lineage tracing model from Garcia et al. showed that mesenchymal cells driven by Hoxb6 and Gli1 expression become pancreatic fibroblasts but only Gli1 positive mesenchymal cells become CAFs in pancreatic ductal adenocarcinoma (PDAC) [8][21]. However, lineage tracing studies have also challenged the belief that most CAFs originate from normal fibroblasts in the tissue—Helms et al. have shown that pancreatic stellate cells (PSCs, the resident fibroblast-like cell of the pancreas) that were lineage traced with Fabp4 only contributed to a fraction of the CAFs in PDAC, contrary to popular belief that most CAFs in PDAC come from PSCs [9][22]. This work demonstrated that there is heterogeneity in the origin of cells that become CAFs in PDAC and potentially in other tumor types as well. Other than lineage tracing, single-cell RNA-sequencing (scRNA-seq) in tandem with either flow-cytometry sorting or mass spectrometry have allowed researchers to identify specific CAF subsets by surface markers that have distinct gene signatures denoting different functions.3.2. Fibroblast Activation
Fibroblasts can be activated into CAFs through several mechanisms. Among growth factors, the TGFβ family ligands are well-established as activating signals [10][34]. Lysophosphatidic acid, a lipid mediator, can also drive expression of the myofibroblast marker αSMA while also increasing the activity of the contractile cytoskeleton [11][35]. Fibroblast activation through Notch signaling [12][13][36,37] has been linked to poor prognosis of ductal breast carcinoma [12][13][36,37]. Several inflammatory cytokines drive CAF subtype differentiation, with IL-6 activating STAT signaling [14][38] and IL-1 acting through NF-κB [15][39]. Stromatogenesis as a result of connective tissue reaction to cancer has been shown to promote desmoplastic fibroblast differentiation in three-dimensional cultures [16][40], and YAP-mediated enhancement of matrix stiffening helps to maintain CAF phenotypes in breast cancer [17][41]. Physiological and genomic stress can also drive fibroblast activation info CAFs, with factors like reactive oxygen species [18][42] and double-stranded DNA breaks [19][43] serving as examples of this phenomenon. There is increasing evidence that cancer cell-derived extracellular vesicles (EVs) can also drive CAF activation [20][21][22][23][25,44,45,46], though EV content differs and can include miRNAs, TGFβ, and cells surface proteins, as well as other proteins, mRNAs, and lncRNAs [21][24][25][44,47,48]. It should be noted, however, that many CAF subtypes and functions exist on a spectrum, rather than distinctive subtypes—and that these subtypes can have overlapping functions, a reality that amplifies the complexity inherent in defining CAFs subtypes.3.3. Fibroblast Markers
CAFs lack a single, specific biomarker in various tumor types. This can be a significant challenge in the context of CAF research. Besides widely mentioned mesenchymal biomarkers such as VIM, αSMA, fibroblast activation protein (FAP), podoplanin (PDPN), integrin beta-1 (ITGB1, also known as CD29), FSP-1 (S100A4), and platelet-derived growth factor alfa (PDGFR-α), PDGFR-β [20][26][7,25], the role of matrix metalloproteinases (MMPs), through which CAFs can facilitate tumor growth, invasion, and metastasis has also been evaluated [27][49]. It is now commonplace for researchers to try to identify useful CAF markers through either flow sorting followed by RNA-seq and/or scRNA-seq to discover new potential CAF markers that are then validated in vivo. The LRRC15 protein represents one example of this—and may also serve as a putative target for precision medicine. High expression of LRRC15 has been reported in CAFs, but not in normal fibroblasts, for multiple tumor types [28][50].4. CAF Subtypes in Various Cancer Types
Flow cytometry and scRNA-seq studies have begun to define the phenotypic and functional heterogeneity of CAFs for individual cancer types, revealing a variety of CAF subtypes (which may also be referred to as “subsets” or “subpopulations”). A cell subtype is defined by specific markers, unique functional properties, and has a secretome pattern that differs from other subtypes. Turley et al. have created a resource for searching CAF and fibroblast markers called “FibroXplorer” [29][52]. It includes data for both human and mouse CAFs/fibroblasts. This is a scRNA-seq object that was created from pooling data from 17 tissues, 50 datasets, and 11 tumor types. Although this may not help with subtyping CAFs in a specific model or a patient sample, it is a great resource for looking at specific CAF markers in the context of various disease states or tissues of origin. Work by the group behind the FibroXplorer resource has also been used to define two universal fibroblast transcriptional sites of origin using mouse tissues, characterize Pi16+ and Col15a1+ expressing fibroblasts, and describe use of dermatopontin (Dpt) to trace fibroblasts [30][53]. While almost all healthy tissue fibroblasts expressed Dpt, especially in Pi16+ and Col15a1+ expressing fibroblasts, Dpt expression has been reported to be lost as a fibroblast becomes specialized either into a CAF or a specific fibroblast niche.
4.1. Breast Cancer
Costa et al., Givel et al., and Bonneau et al. have demonstrated the presence of four CAF subtypes in breast cancer when samples were characterized using multicolor flow cytometry [26][31][32][6,7,57]. The CAF-S1 subset, typically positive for FAP and CD29 and lacking CAV1, contributes to immunosuppression through attracting CD4+ T cells, increasing CD4+CD25+ T lymphocyte survival, and promoting their differentiation into FOXP3+ cells [26][7]. This CAF-S1 subset was preferentially detected in aggressive breast cancer [26][7], in cases of relapsing luminal breast cancer [31][6], and in mesenchymal high-grade serous ovarian cancer associated with poor prognosis [32][57]. On the other hand, the presence of the FAP low CAF-S4 subset in metastatic axillary lymph nodes of breast cancer correlated with the later development of distant metastasis, a finding that suggests that evaluating CAF subtypes may be useful as a prognostic marker for recurrence or metastasis. Compared to the CAF-S1 subset, the CAF-S4 subset expressed lower levels of PDGFRβ but the highest level of CD29. CAF-S2 and CAF-S3 were found not only in tumors but also in healthy tissue, suggesting that they are normal-like resident fibroblasts [31][6]. Using scRNA-seq, Sebastian et al. detected myofibroblast-like CAFs (myCAFs), inflammatory CAFs (iCAFs), and antigen-presenting CAFs (apCAFs) in breast cancer [33][54]. Interestingly, these subsets with similar transcriptomes could also be detected in pancreatic cancer, based on analyses of publicity available RNA-seq data [34][9]. The myCAF and iCAF subtypes were also identified by Wu et al., showing strong enrichment of iCAF signature genes to be associated with cytotoxic T lymphocyte (also known as CD4+ T cells) dysfunction [35][55]. In line with this, patients with a low iCAF dysfunction signature level had a significant survival benefit associated with high cytotoxic T lymphocyte levels [35][55]. MyCAFs, on the other hand, had elevated capabilities for collagen deposition, which has been connected to cancer invasion and disease progress [35][55]. Using immunohistochemistry (IHC), myCAFs were found to be in close proximity to the invasive tumor interface while iCAFs were in an area distal to this interface but with high presence of tumor infiltrating lymphocytes [35][55].4.2. Pancreatic and Gastric Cancer
Pancreatic ductal adenocarcinoma (PDAC) has been the primary subtype of focus of CAF research in pancreatic malignancy, with several authors having delineated CAFs into myCAFs and iCAFs [34][36][37][38][8,9,10,58]. These two subpopulations of CAFs have been identified using immunofluorescence (IF), IHC, and microscopy [37][10]. Using scRNA-seq, 962 fibroblast cells could be analyzed in vivo and two distinct subclusters could be formed with unique gene signatures [34][9]. The subcluster 1 had enriched expression of Il-6 and Il-8 and chemokines such as Cxcl1, Cxcl2, Ccl2, and Cxcl12, and was identified as iCAFs. The IL-6 expressing iCAFs were located far away from neoplastic cells in the desmoplastic stroma according to IHC [37][10]. The αSMA positive subcluster 2 was identified as myCAFs and spatially distributed in direct proximity to neoplastic cells forming a periglandular ring surrounding the clusters of cancer cells [34][37][9,10]. Using Gene Set Enrichment Analysis (GSEA), enriched pathways could be identified enclosing an upregulation of inflammatory pathways such as IFNγ response, TNF/NF-κB, IL2/STAT5, IL6/JAK/STAT3, and the complement pathway in iCAFs, while myCAFs displayed an upregulation in pathways such as smooth muscle contraction, focal adhesion, ECM organization and collagen formation. Biffi et al. further revealed IL1 to be critical for generation of iCAFs [36][8]. When analyzing PDAC tumors in Kras+/LSL-G12D; Trp53+/LSL-R172H; Pdx1-Cre (KPC) mice, a new subpopulation—namely, apCAF—were described [34][9]. These MHC II-expressing and Cd74-expressing apCAFs could activate CD4+ T cells in an antigen-specific fashion based on use of an ovalbumin-specific TCR transgenic OTII mouse model. An apCAF subtype has also been suggested by Zhang et al. in HNSCC [39][62]. Another group also divided PDAC CAFs into myCAFs and iCAFs, with IL1 inducing an iCAF phenotype similar to the one reported by Biffi et al. [40][59]. However, this other group also revealed that hypoxia promoted the iCAF-like state. The TME is characterized by varied extent of hypoxia and associated with poor prognosis of patients with PDAC. This finding links the direct functional impact of hypoxia on CAFs in the TME. A third group also delineated CAFs into myCAFs and iCAFs [41][60]. They showed in an orthotopic model of diffuse-type gastric cancer (DGC) that EVs from highly metastatic DGC cells transferred various miRNAs and induced chemokine expression (CXCL1 and CXCL8) in fibroblasts.4.3. Hepatocellular Carcinoma and Cholangiocarcinoma
Ying et al. reviewed studies summarizing CAF subpopulations and their markers in hepatocellular carcinoma (HCC) and cholangiocarcinoma (CCA), highlighting the problematic lack of specific CAF markers [42][43][11,12]. Six distinct fibroblast subsets were detected using scRNA-seq on human intrahepatic cholangiocarcinoma (ICC) by Zhang et al.: vascular CAFs (vCAF), matrix CAFs (mCAF), inflammatory CAFs (iCAF), antigen-presenting CAFs (apCAF), EMT-like CAFs (eCAF), and lipofibroblasts. The authors determined that the vCAF subgroup was the most prevalent in the tumors. These vCAFs were shown to secrete IL-6, which induced significant epigenetic alterations in tumor cells and furthered malignancy [44][13].5. CAF Subtypes in OSCC
Given the emerging complexity and varied reports about CAF subtypes, it is clear that resolving the role of these cells in cancer processes will remain challenging for the near future. It is tempting to suggest that CAF subtypes described by one group are the same as subtypes described by another group with the only difference being the name. F While several research groups have detected and categorized CAF subtypes in breast, ovarian, pancreatic, and hepatocellular cancer, the work to describe CAF subtypes in OSCC has just begun. Figure 1 illustrates CAF subtypes identified in several cancer types including OSCC. Galbo et al. attempted to find common ground between CAF subgroupings by analyzing results from head and neck squamous cell carcinoma (HNSCC, most frequently presented as OSCC), melanoma, and lung cancer (LC) [45][67].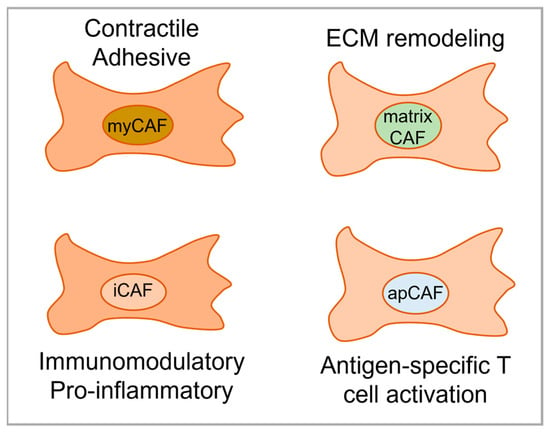
6. The Prognostic Role of CAF Subtypes in OSCC
7. Therapeutic Opportunities Linked to CAF Subtypes
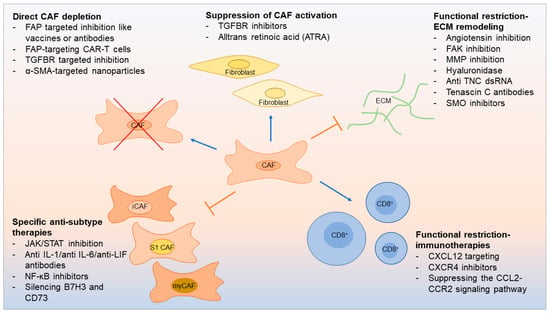
In PDAC, up to 90% of the tumor volume consists of stroma [40][59]. In HNSCC, CAFs can account for up to 80% of the tumor mass [48][72]. The tumor–stroma ratio has been shown to have prognostic importance [49][73], and evaluating this ratio can also garner insights into clinical disease [50][74]. Tumor stroma may block delivery of therapy to cancer cells, support cancer growth, and promote therapy resistance [25][48]. On the other hand, the tumor stroma may also contain potential targets for potential therapy. Recently, the number of preclinical experiments targeting CAFs to restore the anticancer immune response has increased dramatically. Further, strategies for CAF-based immunotherapy have emerged: direct CAF depletion including a novel therapeutic that uses FAP-targeting chimeric antigen receptor (CAR)-T cells [5] resulting in reduced CAF activation [51][75] and functional suppression of CAF-induced ECM remodeling [52][53][54][76,77,78] (Figure 2).