α-linolenic acid (ALA) belongs to the family of n-3 polyunsaturated fatty acids (n-3 PUFAs) and contains a carbon–carbon double bond on the third carbon atom at the methyl end of the carbon chain. This family of essential fatty acids also includes eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA). ALA has gradually attracted increased attention due to its nutritional and medicinal advantages. Studies have shown that ALA exerts beneficial effects on a variety of diseases, including cancer.
- α-linolenic acid
- anticancer
- cell proliferation
- apoptosis
- inflammatory response
- tumor metastasis
- antioxidant
1. Introduction
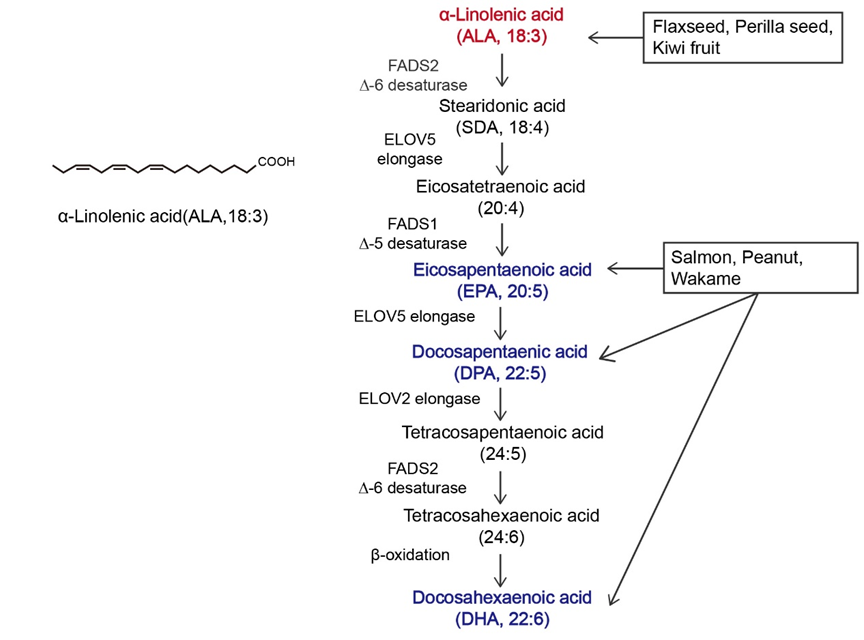
Figure 1.Overview of α-linolenic acid (ALA). Molecular structure of ALA and the in vivo metabolic pathway by which n-3 polyunsaturated fatty acids (n-3 PUFAs) are generated from ALA. The colors in the figure are intended to emphasize the members of n-3 PUFAs. The vertical arrow represents the metabolism of ALA to other n-3 PUFAs in vivo; Arrows in other directions represent different dietary sources of n-3 PUFAs.
According to the International Agency for Research on Cancer GLOBOCAN 2020 cancer incidence and mortality estimates, in 2020, there were 19.3 million new cancer cases and nearly 10 million deaths worldwide [30], except for melanoma cell cancer. As a populous country, China’s new cancer cases in 2020 accounted for 24% of the world’s new cancer cases [31]. Cancer has surpassed cardiovascular disease as the leading cause of death in China. A prominent feature of tumors is that their growth and proliferation are uncontrolled, and invasion and metastasis are the main problems facing current cancer treatment. Although the etiology of cancer is not yet fully understood, it can be roughly divided into two categories, endogenous and exogenous, and nutrients are the factors most closely related to daily life [32]. Nutrient intake can regulate the tumor microenvironment, thereby affecting cancer cell proliferation, apoptosis, and invasion. Current cancer treatment strategies, including surgery, radiotherapy, and chemotherapy, reduce the quality of life of patients, and diet has gradually become one of the most common treatment methods due to its high acceptance by patients and low toxicity and side effects [13][14]. Most of the initial dietary studies focused on limiting the proliferation of tumor cells by reducing the supply of major nutrients to tumors [33][34][35]. With further research, supplementation with specific nutrients, including histidine and mannose, has also become a strategy for the clinical treatment of cancer [36][37].The n-3 PUFA family has attracted considerable attention for its anticancer effects and use as a dietary supplement.
Cancer has been a constant threat to human threat to human life since its identification, and even when it is treatable, it greatly reduces quality of life. Many studies have shown that ALA exerts significant anticancer effects on multiple cancers [38][39][40][41][42][43][44][45][46][47][48][38,39,40,41,42,43,44,45,46,47,48]. In Table 1, a subset of ALA-sensitive cancers is listed, including prostate cancer, BC, hepatocellular carcinoma, colorectal cancer (CRC), and pancreatic cancer. In addition, ALA also exerts effects on many common gastrointestinal tumors and bladder cancer [49][50][51][49,50,51]. As shown in Figure 21, ALA exerts a variety of anticancer effects, including inhibiting proliferation, inducing apoptosis, suppressing tumor metastasis and angiogenesis, and exerting antioxidant effects. To provide a brief introduction to the anticancer effects of ALA, these effects are systematically reviewed, focusing on pharmacological actions and molecular mechanisms.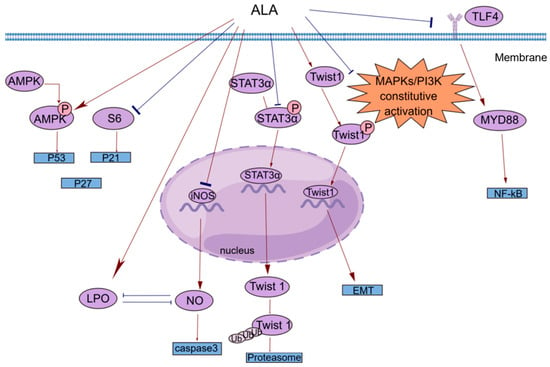
| Cancer | Effect | Effector Molecules | Change in Ex-Pression |
|---|---|---|---|
| PCa (prostate cancer) [52] |
anti-inflammatory effect | PG/LTs | downregulation |
| BC (breast cancer) [38][39][38,39] |
anti-inflammatory effect/inhibition of tumor metastasis | COX2/PGE2/Twist 1 | downregulation |
| HCC (hepatocellular carcinoma) [40][41][40,41] |
inhibition of proliferation | Farnesoid X receptor | upregulation |
| CRC (colorectal cancer) [42][43][42,43] |
induction of apoptosis | caspase 3 | downregulation |
| PCA (pancreatic cancer) [44] |
anti-inflammatory effect | IL-1β/IL-6 | downregulation |