Heat stress represents a pervasive global concern with far-reaching implications for the reproductive efficiency of both animal and human populations. An extensive body of published research on heat stress effects utilizes controlled experimental environments to expose cells and tissues to heat stress and its disruptive influence on the physiological aspects of reproductive phenotypic traits, encompassing parameters such as sperm quality, sperm motility, viability, and overall competence. Beyond these immediate effects, heat stress has been linked to embryo losses, compromised oocyte development, and even infertility across diverse species. One of the primary mechanisms underlying these adverse reproductive outcomes is the elevation of reactive oxygen species (ROS) levels precipitating oxidative stress and apoptosis within mammalian reproductive cells. Oxidative stress and apoptosis are recognized as pivotal biological factors through which heat stress exerts its disruptive impact on both male and female reproductive cells. In a concerted effort to mitigate the detrimental consequences of heat stress, supplementation with antioxidants, both in natural and synthetic forms, has been explored as a potential intervention strategy. Furthermore, reproductive cells possess inherent self-protective mechanisms that come into play during episodes of heat stress, aiding in their survival.
- heat stress
- oxidative stress
- apoptosis
- antioxidants
- mammalian reproductive cells
- fertility
1. Introduction
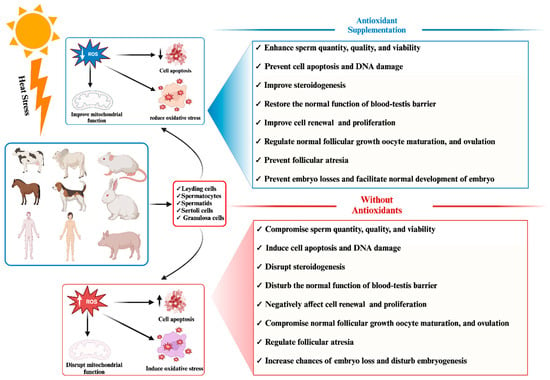
2. Impact of Heat-Stress-Induced Oxidative Stress and Apoptosis on Mammalian Reproductive Cell Functionality
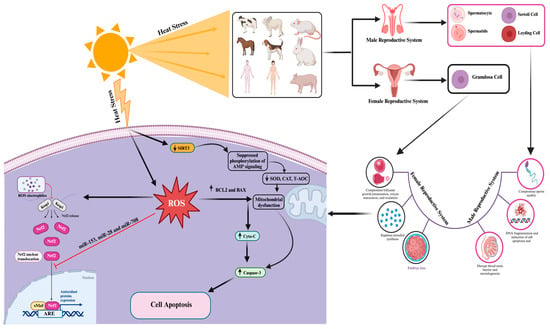
 ” indicates the effect has been suppressed.
” indicates the effect has been suppressed.| Heat Stress | Biological Effect | Cells | Reference |
|---|---|---|---|
|
Sertoli cells | [116] | |
|
Sertoli cells | [106] | |
|
Testis | [117] | |
|
Ovarian granulosa cells | [95] | |
|
Mouse granulosa cells | [118] |
3. Advancement and Understanding of Genetic Biomarkers Associated with Heat Stress Resistance and Reduced Apoptosis and Oxidative Stress in Mammalian Reproductive Cells
3.1. Role of Heat Shock Protein (HSP) Genes in Mitigating Heat-Stress-Induced Oxidative Stress and Apoptosis in Mammalian Reproductive Cells
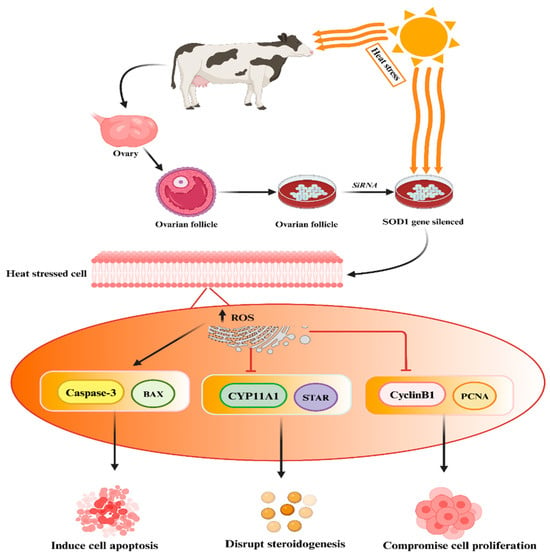
3.2. Protective Role of SOD Genes against Heat-Stress-Induced Oxidative Stress and Apoptosis in Mammalian Reproductive Cells
 ” indicates the effect has been suppressed.
” indicates the effect has been suppressed.3.3. ERK1/2 Signaling Pathway Protects Mammalian Reproductive Cells from Heat-Stress-Induced Apoptosis
3.4. Protective Role of Nrf2 in Protection of Mammalian Cells against Heat-Stress-Induced Oxidative Stress and Apoptosis
3.5. Role of Adenosine 5′-Monophosphate-Activated Protein Kinase (AMPK) in Self-Recovery from Heat-Stress-Induced Oxidative Stress and Apoptosis
References
- Becker, C.A.; Collier, R.J.; Stone, A.E. Invited review: Physiological and behavioral effects of heat stress in dairy cows. J. Dairy Sci. 2020, 103, 6751–6770.
- Schulte, P.A.; Bhattacharya, A.; Butler, C.R.; Chun, H.K.; Jacklitsch, B.; Jacobs, T.; Kiefer, M.; Lincoln, J.; Pendergrass, S.; Shire, J.; et al. Advancing the framework for considering the effects of climate change on worker safety and health. J. Occup. Environ. Hyg. 2016, 13, 847–865.
- Ruane, A.C.; Vautard, R.; Ranasinghe, R.; Sillmann, J.; Coppola, E.; Arnell, N.; Cruz, F.A.; Dessai, S.; Iles, C.E.; Islam, A.K.M.S.; et al. The Climatic Impact-Driver Framework for Assessment of Risk-Relevant Climate Information. Earths Future 2022, 10, e2022EF002803.
- Shibasaki, M.; Crandall, C.G. Mechanisms and controllers of eccrine sweating in humans. Front. Biosci. 2010, 2, 685–696.
- Hanna, E.G.; Tait, P.W. Limitations to Thermoregulation and Acclimatization Challenge Human Adaptation to Global Warming. Int. J. Environ. Res. Public Health 2015, 12, 8034–8074.
- Thompson, V.; Mitchell, D.; Hegerl, G.C.; Collins, M.; Leach, N.J.; Slingo, J.M. The most at-risk regions in the world for high-impact heatwaves. Nat. Commun. 2023, 14, 2152.
- Walter, E.J.; Carraretto, M. The neurological and cognitive consequences of hyperthermia. Crit. Care 2016, 20, 199.
- Zhang, G.; Han, L.; Yao, J.; Yang, J.; Xu, Z.; Cai, X.; Huang, J.; Pei, L. Assessing future heat stress across China: Combined effects of heat and relative humidity on mortality. Front. Public Health 2023, 11, 1282497.
- Dovolou, E.; Giannoulis, T.; Nanas, I.; Amiridis, G.S. Heat Stress: A Serious Disruptor of the Reproductive Physiology of Dairy Cows. Animals 2023, 13, 1846.
- Tirpák, F.; Halo, M.; Tomka, M.; Slanina, T.; Tokárová, K.; Błaszczyk-Altman, M.; Dianová, L.; Ivanič, P.; Kirchner, R.; Greń, A.; et al. Sperm Quality Affected by Naturally Occurring Chemical Elements in Bull Seminal Plasma. Antioxidants 2022, 11, 1796.
- Tirpák, F.; Greifová, H.; Lukáč, N.; Stawarz, R.; Massányi, P. Exogenous factors affecting the functional integrity of male reproduction. Life 2021, 11, 213.
- Boni, R. Heat stress, a serious threat to reproductive function in animals and humans. Mol. Reprod. Dev. 2019, 86, 1307–1323.
- Cong, X.; Zhang, Q.; Li, H.; Jiang, Z.; Cao, R.; Gao, S.; Tian, W. Puerarin ameliorates heat stress–induced oxidative damage and apoptosis in bovine Sertoli cells by suppressing ROS production and upregulating Hsp72 expression. Theriogenology 2017, 88, 215–227.
- Cao, K.X.; Deng, Z.C.; Liu, M.; Huang, Y.X.; Yang, J.C.; Sun, L.H. Heat Stress Impairs Male Reproductive System with Potential Disruption of Retinol Metabolism and Microbial Balance in the Testis of Mice. J. Nutr. 2023, 153, 3373–3381.
- Costa, G.M.; Lacerda, S.M.; Figueiredo, A.F.; Leal, M.C.; Rezende-Neto, J.V.; França, L.R. Higher Environmental Temperatures Promote Acceleration of Spermatogenesis in Vivo in Mice (Mus musculus). J. Therm. Biol. 2018, 77, 14–23.
- Huang, D.; Cai, J.; Zhang, C.; Jin, R.; Bai, S.; Yao, F.; Ding, H.; Zhao, B.; Chen, Y.; Wu, X.; et al. Semen quality and seminal plasma metabolites in male rabbits (Oryctolagus cuniculus) under heat stress. PeerJ 2023, 11, e15112.
- Lyrio, L.L.; Lazaro, M.A.; Sonegheti, R.; Moulin, L.; Coslop, L.; Sarto, C.G.; Loureiro, B.; Favoreto, M.G. Effects of heat stress on sperm quality of French Bulldogs. Theriogenology 2023, 199, 131–137.
- Takalani, N.B.; Monageng, E.M.; Mohlala, K.; Monsees, T.K.; Henkel, R.; Opuwari, C.S. Role of oxidative stress in male infertility. Reprod. Fertil. 2023, 4, 3.
- Netherton, J.K.; Robinson, B.R.; Ogle, R.A.; Gunn, A.; Villaverde, A.I.; Colyvas, K.; Wise, C.; Russo, T.; Dowdell, A.; Baker, M.A. Seasonal variation in bull semen quality demonstrates there are heat-sensitive and heat-tolerant bulls. Sci. Rep. 2022, 12, 15322.
- Morrell, J.M. Heat stress and bull fertility. Theriogenology 2020, 153, 62–67.
- Sabés-Alsina, M.; Wallgren, M.; Sjunnesson, Y.C.; Ntallaris, T.; Lundeheim, N.; López-Béjar, M.; Morrell, J.M. Effect of season on the in vitro fertilizing ability of frozen–thawed Spanish bovine spermatozoa. J. Dairy Sci. 2020, 103, 9525–9533.
- Shahat, A.M.; Rizzoto, G.; Kastelic, J.P. Amelioration of heat stress-induced damage to testes and sperm quality. Theriogenology 2020, 158, 84–96.
- Wrzecińska, M.; Kowalczyk, A.; Kordan, W.; Cwynar, P.; Czerniawska-Piątkowska, E. Disorder of Biological Quality and Autophagy Process in Bovine Oocytes Exposed to Heat Stress and the Effectiveness of In Vitro Fertilization. Int. J. Mol. Sci. 2023, 24, 11164.
- Capela, L.; Leites, I.; Romao, R.; Lopes-da-Costa, L.; Pereira, R. Impact of Heat Stress on Bovine Sperm Quality and Competence. Animals 2022, 12, 975.
- Llamas-Luceño, N.; Hostens, M.; Mullaart, E.; Broekhuijse, M.; Lonergan, P.; Van Soom, A. High temperature-humidity index compromises sperm quality and fertility of Holstein bulls in temperate climates. J. Dairy Sci. 2020, 103, 9502–9514.
- Petrocchi Jasinski, F.; Evangelista, C.; Basiricò, L.; Bernabucci, U. Responses of Dairy Buffalo to Heat Stress Conditions and Mitigation Strategies: A Review. Animals 2023, 13, 1260.
- Papa, P.M.; Segabinazzi, L.G.; Fonseca-Alves, C.E.; Papa, F.O.; Alvarenga, M.A. Intratesticular transplantation of allogenic mesenchymal stem cells mitigates testicular destruction after induced heat stress in Miniature-horse stallions. J. Equine Vet. Sci. 2023, 132, 104961.
- Shakeel, M.; Yoon, M. Heat stress and stallion fertility. J. Anim. Sci. Technol. 2023, 65, 683–697.
- Shakeel, M.; Yoon, M. Effects of insulin-like growth factor-1 on the proliferation and apoptosis of stallion testicular cells under normal and heat stress culture conditions. Anim. Reprod. Sci. 2023, 256, 107319.
- Ebeid, T.A.; Aljabeili, H.S.; Al-Homidan, I.H.; Volek, Z.; Barakat, H. Ramifications of heat stress on rabbit production and role of nutraceuticals in alleviating its negative impacts: An updated review. Antioxidants 2023, 12, 1407.
- de Andrade, A.F.; Balogun, K.; Machaty, Z.; Knox, R.V. Effects of supplemental antioxidants on in vitro fertility measures for cryopreserved boar spermatozoa. Theriogenology 2023, 200, 33–42.
- Abedin, S.N.; Baruah, A.; Baruah, K.K.; Bora, A.; Dutta, D.J.; Kadirvel, G.; Katiyar, R.; Doley, S.; Das, S.; Khargharia, G.; et al. Zinc oxide and selenium nanoparticles can improve semen quality and heat shock protein expression in cryopreserved goat (Capra hircus) spermatozoa. J. Trace Elem. Med. Biol. 2023, 80, 127296.
- Habibi, P.; Ostad, S.N.; Heydari, A.; Monazzam, M.R.; Foroushani, A.R.; Ghazi-Khansari, M.; Golbabaei, F. Diagnostic biomarkers of heat stress induced-DNA in occupational exposure: A systematic review. J. Health Saf. Work 2023, 12, 800–819.
- Robinson, B.R.; Netherton, J.K.; Ogle, R.A.; Baker, M.A. Testicular heat stress, a historical perspective and two postulates for why male germ cells are heat sensitive. Biol. Rev. 2023, 98, 603–622.
- Abd El-Emam, M.M.; Ray, M.N.; Ozono, M.; Kogure, K. Heat stress disrupts spermatogenesis via modulation of sperm-specific calcium channels in rats. J. Therm. Biol. 2023, 112, 103465.
- Gan, M.; Jing, Y.; Xie, Z.; Ma, J.; Chen, L.; Zhang, S.; Zhao, Y.; Niu, L.; Wang, Y.; Li, X.; et al. Potential Function of Testicular MicroRNAs in Heat-Stress-Induced Spermatogenesis Disorders. Int. J. Mol. Sci. 2023, 24, 8809.
- Gao, W.J.; Li, H.X.; Feng, J.; Lu, X.R.; Yin, P.L.; Jia, H.; Ma, W.Z. Transcriptome Analysis in High Temperature Inhibiting Spermatogonial Stem Cell Differentiation In Vitro. Reprod. Sci. 2023, 30, 1938–1951.
- Hu, Y.; Li, Q.; Qian, Z.; Luo, K.; Luo, N. Joint Analysis of Genome-wide DNA Methylation and Transcription Sequencing Identifies the Role of BAX Gene in Heat Stress–Induced-Sertoli Cells Apoptosis. Reprod. Sci. 2024, 1–12.
- Liu, C.X.; Hu, S.Q.; Liu, D.L.; Xu, Y.H.; Hu, K.; Guo, J. The effect of semen cuscutae flavonoid on Sertoli cells and blood-testis barrier in male infertility: Integrating network pharmacology and experimental verification. Pharm. Biol. 2023, 61, 986–999.
- Wang, Y.; Wu, Z.W.; Mou, Q.; Chen, L.; Fang, T.; Zhang, Y.Q.; Yin, Z.; Du, Z.Q.; Yang, C.X. Global 3′-UTRome of porcine immature Sertoli cells altered by acute heat stress. Theriogenology 2023, 196, 79–87.
- Yang, J.; Ou, X.; Shu, M.; Wang, J.; Zhang, X.; Wu, Z.; Hao, W.; Zeng, H.; Shao, L. Inhibition of p38MAPK Signaling Pathway Alleviates Radiation-Induced Testicular Damage through Improving Spermatogenesis. Brit. J. Pharmacol. 2024, 181, 393–412.
- Zhong, L.; Zhong, H.; Dai, Y.; Zhang, Q. Effect of heat shock transcription factor 5 (Hsf5) knockdown on heat shock family in mouse Leydig cells and Sertoli cells. Basic Clin. Med. 2023, 43, 626.
- Deng, X.; Wang, Q.; Shi, C.; Wei, J.; Lv, Z.; Huang, S.; Duan, Y.G.; Zhang, X.; Liu, Y. Heat wave exposure and semen quality in sperm donation volunteers: A retrospective longitudinal study in south China. Environ. Res. 2023, 236, 116665.
- Mai, H.; Ke, J.; Li, M.; He, M.; Qu, Y.; Jiang, F.; Cai, S.; Xu, Y.; Fu, L.; Pi, L.; et al. Association of living environmental and occupational factors with semen quality in Chinese men: A cross-sectional study. Sci. Rep. 2023, 13, 15671.
- Tian, R.; Yang, T.; Xiao, C.; Li, F.; Fu, L.; Zhang, L.; Cai, J.; Zeng, S.; Liao, J.; Song, G.; et al. Outdoor artificial light at night and male sperm quality: A retrospective cohort study in China. Environ. Pollut. 2023, 341, 122927.
- Zhang, X.; Fan, Z.; Wang, Q.; Deng, X.; Xu, R.; Li, Y.; Liu, T.; Wang, R.; Shi, C.; Huang, S.; et al. Association between ambient temperature and semen quality among sperm donation volunteers in South China. Environ. Int. 2023, 173, 107809.
- Khan, I.; Mesalam, A.; Heo, Y.S.; Lee, S.H.; Nabi, G.; Kong, I.K. Heat Stress as a Barrier to Successful Reproduction and Potential Alleviation Strategies in Cattle. Animals 2023, 13, 2359.
- St-Pierre, N.R.; Cobanov, B.; Schnitkey, G. Economic losses from heat stress by US livestock industries. J. Dairy Sci. 2003, 86, E52–E77.
- Rhoads, M.L. Reproductive consequences of whole-body adaptations of dairy cattle to heat stress. Animal 2023, 17, 100847.
- Stefanska, B.; Sobolewska, P.; Fievez, V.; Pruszynska-Oszmałek, E.; Purwin, C.; Nowak, W. The impact of heat stress on performance, fertility, and adipokines involved in regulating systemic immune response during lipolysis of early lactating dairy cows. J. Dairy Sci. 2023.
- Barragán, A.L.; Avendaño-Reyes, L.; Mellado-Bosque, M.; Meza-Herrera, C.A.; Vicente-Pérez, R.; Castañeda, V.J.; Díaz-Molina, R.; Macías-Cruz, U. Seasonal heat stress compromises testicular thermoregulation and semen quality of Dorper rams raised in a desert climate. J. Therm. Biol. 2023, 118, 103737.
- Negrón-Pérez, V.M.; Fausnacht, D.W.; Rhoads, M.L. Invited review: Management strategies capable of improving the reproductive performance of heat-stressed dairy cattle. J. Dairy Sci. 2019, 102, 10695–10710.
- Rahman, M.B.; Schellander, K.; Luceno, N.L.; Van Soom, A. Heat stress responses in spermatozoa: Mechanisms and consequences for cattle fertility. Theriogenology 2018, 113, 102–112.
- de Aguiar, L.H.; Hyde, K.A.; Pedroza, G.H.; Denicol, A.C. Heat stress impairs in vitro development of preantral follicles of cattle. Anim. Reprod. Sci. 2020, 213, 106277.
- Khan, A.; Dou, J.; Wang, Y.; Jiang, X.; Khan, M.Z.; Luo, H.; Usman, T.; Zhu, H. Evaluation of heat stress effects on cellular and transcriptional adaptation of bovine granulosa cells. J. Anim. Sci. Biotechnol. 2020, 11, 25.
- Alemu, T.W.; Pandey, H.O.; Wondim, D.S.; Gebremedhn, S.; Neuhof, C.; Tholen, E.; Holker, M.; Schellander, K.; Tesfaye, D. Oxidative and endoplasmic reticulum stress defense mechanisms of bovine granulosa cells exposed to heat stress. Theriogenology 2018, 110, 130–141.
- De Rensis, F.; Lopez-Gatius, F.; García-Ispierto, I.; Morini, G.; Scaramuzzi, R.J. Causes of declining fertility in dairy cows during the warm season. Theriogenology 2017, 91, 145–153.
- Schüller, L.K.; Michaelis, I.; Heuwieser, W. Impact of heat stress on estrus expression and follicle size in estrus under field conditions in dairy cows. Theriogenology 2017, 102, 48–53.
- Ho, K.T.; Balboula, A.Z.; Homma, K.; Takanari, J.; Bai, H.; Kawahara, M.; Nguyen, K.T.; Takahashi, M. Synergistic effect of standardized extract of Asparagus officinalis stem and heat shock on progesterone synthesis with lipid droplets and mitochondrial function in bovine granulosa cells. J. Steroid Biochem. Mol. Biol. 2023, 225, 106181.
- Menjivar, N.G.; Gad, A.; Gebremedhn, S.; Ghosh, S.; Tesfaye, D. Granulosa cell-derived extracellular vesicles mitigate the detrimental impact of thermal stress on bovine oocytes and embryos. Front. Cell Dev. Biol. 2023, 11, 1142629.
- Miękiewska, K.; Kordowitzki, P.; Pareek, C.S. Effects of Heat Stress on Bovine Oocytes and Early Embryonic Development—An Update. Cells 2022, 11, 4073.
- Yüzen, D.; Graf, I.; Diemert, A.; Arck, P.C. Climate change and pregnancy complications: From hormones to the immune response. Front. Endocrinol. 2023, 14, 1149284.
- Chotimanukul, S.; Suwimonteerabutr, J.; Techakumphu, M.; Swangchan-Uthai, T. In vitro effects of short-term and long-term heat exposures on the immune response and prostaglandin biosynthesis in bovine endometrial cells. Animals 2022, 12, 2359.
- Roth, Z. Heat stress reduces maturation and developmental capacity in bovine oocytes. Reprod. Fertil. Dev. 2021, 33, 66–75.
- Roth, Z. Influence of heat stress on reproduction in dairy cows—Physiological and practical aspects. J. Anim. Sci. 2020, 98, S80–S87.
- Roth, Z. Reproductive physiology and endocrinology responses of cows exposed to environmental heat stress-Experiences from the past and lessons for the present. Theriogenology 2020, 155, 150–156.
- Huber, E.; Notaro, U.S.; Recce, S.; Rodríguez, F.M.; Ortega, H.H.; Salvetti, N.R.; Rey, F. Fetal programming in dairy cows: Effect of heat stress on progeny fertility and associations with the hypothalamic-pituitary-adrenal axis functions. Anim. Reprod. Sci. 2020, 216, 106348.
- Nanas, I.; Chouzouris, T.M.; Dadouli, K.; Dovolou, E.; Stamperna, K.; Barbagianni, M.; Valasi, I.; Tsiaras, A.; Amiridis, G.S. A study on stress response and fertility parameters in phenotypically thermotolerant and thermosensitive dairy cows during summer heat stress. Reprod. Domest. Anim. 2020, 55, 1774–1783.
- Payton, R.R.; Rispoli, L.A.; Nagle, K.A.; Gondro, C.; Saxton, A.M.; Voy, B.H.; Edwards, J.L. Mitochondrial-related consequences of heat stress exposure during bovine oocyte maturation persist in early embryo development. J. Reprod. Dev. 2018, 64, 243–251.
- Diaz, F.A.; Gutierrez-Castillo, E.J.; Foster, B.A.; Hardin, P.T.; Bondioli, K.R.; Jiang, Z. Evaluation of seasonal heat stress on transcriptomic profiles and global DNA methylation of bovine oocytes. Front. Genet. 2021, 12, 699920.
- Gendelman, M.; Roth, Z. In vivo vs. in vitro models for studying the effects of elevated temperature on the GV-stage oocyte, subsequent developmental competence and gene expression. Anim. Reprod. Sci. 2012, 134, 125–134.
- Gendelman, M.; Roth, Z. Incorporation of coenzyme Q10 into bovine oocytes improves mitochondrial features and alleviates the effects of summer thermal stress on developmental competence. Biol. Reprod. 2012, 87, 118.
- Makris, A.; Alevra, A.I.; Exadactylos, A.; Papadopoulos, S. The Role of Melatonin to Ameliorate Oxidative Stress in Sperm Cells. Int. J. Mol. Sci. 2023, 24, 15056.
- Zhong, R.Z.; Zhou, D.W. Oxidative Stress and Role of Natural Plant-Derived Antioxidants in Animal Reproduction. J. Integr. Agric. 2013, 12, 1826–1838.
- Mohlala, K.; Offor, U.; Monageng, E.; Takalani, N.B.; Opuwari, C.S. Overview of the Effects of Moringa oleifera Leaf Extract on Oxidative Stress and Male Infertility: A Review. Appl. Sci. 2023, 13, 4387.
- Shahat, A.M.; Thundathil, J.C.; Kastelic, J.P. Melatonin improves post-thaw sperm quality after mild testicular heat stress in rams. Reprod. Domest. Anim. 2023, 58, 423–430.
- El-Gindy, Y.M.; Sabir, S.A.; Zahran, S.M.; Morshedy, S.A. The protective effect of aqueous orange peel extract against severe heat stress on reproductive efficiency, milk yield, and antioxidant status of female rabbits. J. Therm. Biol. 2023, 111, 103403.
- El-Gindy, Y.M.; Sabir, S.A.; Zahran, S.M.; Ahmed, M.H.; Reuben, R.C.; Salem, A.Z. Effect of dietary onion (Allium cepa L.) powder as an antioxidant on semen quality, blood biochemicals, and reproductive parameters, as well as immunological variables of rabbit bucks under severe heat stress. Trop. Anim. Health Prod. 2023, 55, 380.
- El-Gindy, Y.M.; Zahran, S.M.; Ahmed, M.H.; Adegbeye, M.J.; Salem, A.Z.; Salam, M.Y. Enhancing semen quality, antioxidant status and sex hormones of V-line rabbit bucks fed on supplemented diets with dried moringa leaves. Anim. Biotechnol. 2023, 34, 2626–2635.
- El-Ratel, I.T.; El-Kholy, K.H.; Mousa, N.A.; El-Said, E.A. Impacts of selenium nanoparticles and spirulina alga to alleviate the deleterious effects of heat stress on reproductive efficiency, oxidative capacity and immunity of doe rabbits. Anim. Biotechnol. 2023, 34, 3519–3532.
- Jimoh, O.A.; Daramola, O.T.; Okin-Aminu, H.O.; Ojo, O.A.; Oyeyemi, W.A. Effect of phytogenic supplements on the reproductive physiology and metabolic hormones of rabbits exposed to heat stress conditions. J. Therm. Biol. 2023, 112, 103438.
- Naspinska, R.; Moreira da Silva, M.H.; Moreira da Silva, F. Current Advances in Bovine In Vitro Maturation and Embryo Production Using Different Antioxidants: A Review. J. Dev. Biol. 2023, 11, 36.
- Toosinia, S.; Davoodian, N.; Arabi, M.; Kadivar, A. Ameliorating Effect of Sodium Selenite on Developmental and Molecular Response of Bovine Cumulus-Oocyte Complexes Matured In Vitro under Heat Stress Condition. Biol. Trace Elem. Res. 2024, 202, 161–174.
- Tripathi, S.K.; Nandi, S.; Gupta, P.S.; Mondal, S. Antioxidants supplementation improves the quality of in vitro produced ovine embryos with amendments in key development gene expressions. Theriogenology 2023, 201, 41–52.
- Yaacobi-Artzi, S.; Shimoni, C.; Kalo, D.; Hansen, P.J.; Roth, Z. Melatonin slightly alleviates the effect of heat shock on bovine oocytes and resulting blastocysts. Theriogenology 2020, 158, 477–489.
- Rakha, S.I.; Elmetwally, M.A.; El-Sheikh Ali, H.; Balboula, A.; Mahmoud, A.M.; Zaabel, S.M. Importance of Antioxidant Supplementation during In Vitro Maturation of Mammalian Oocytes. Vet. Sci. 2022, 9, 439.
- Zhang, B.; Pan, C.; Feng, C.; Yan, C.; Yu, Y.; Chen, Z.; Guo, C.; Wang, X. Role of mitochondrial reactive oxygen species in homeostasis regulation. Redox Rep. 2022, 27, 45–52.
- Aitken, R.J.; Drevet, J.R.; Moazamian, A.; Gharagozloo, P. Male infertility and oxidative stress: A focus on the underlying mechanisms. Antioxidants 2022, 11, 306.
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The chemistry of reactive oxygen species (ROS) revisited: Outlining their role in biological macromolecules (DNA, lipids and proteins) and induced pathologies. Int. J. Mol. Sci. 2021, 22, 4642.
- Belhadj Slimen, I.; Najar, T.; Ghram, A.; Dabbebi, H.; Ben Mrad, M.; Abdrabbah, M. Reactive oxygen species, heat stress and oxidative-induced mitochondrial damage. A review. Int. J. Hyperth. 2014, 30, 513–523.
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative stress and male infertility. Nat. Rev. Urol. 2017, 14, 470–485.
- Drevet, J.R.; Hallak, J.; Nasr-Esfahani, M.H.; Aitken, R.J. Reactive oxygen species and their consequences on the structure and function of mammalian spermatozoa. Antioxid. Redox Signal. 2022, 37, 481–500.
- Sharma, P.; Kaushal, N.; Saleth, L.R.; Ghavami, S.; Dhingra, S.; Kaur, P. Oxidative stress-induced apoptosis and autophagy: Balancing the contrary forces in spermatogenesis. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2023, 1869, 166742.
- Aldahhan, R.A.; Stanton, P.G.; Ludlow, H.; de Kretser, D.M.; Hedger, M.P. Acute heat-treatment disrupts inhibin-related protein production and gene expression in the adult rat testis. Mol. Cell. Endocrinol. 2019, 498, 110546.
- Khan, A.; Khan, M.Z.; Dou, J.; Xu, H.; Liu, L.; Zhu, H.; Wang, Y. SOD1 Gene silencing promotes apoptosis and suppresses proliferation of heat-stressed bovine granulosa cells via induction of oxidative stress. Vet. Sci. 2021, 8, 326.
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 2015, 13, 37.
- Henriques, M.C.; Santiago, J.; Patrício, A.; Herdeiro, M.T.; Loureiro, S.; Fardilha, M. Smoking Induces a Decline in Semen Quality and the Activation of Stress Response Pathways in Sperm. Antioxidants 2023, 12, 1828.
- Li, H.; Cong, X.; Yu, W.; Jiang, Z.; Fu, K.; Cao, R.; Tian, W.; Feng, Y. Baicalin inhibits oxidative injures of mouse uterine tissue induced by acute heat stress through activating the Keap1/Nrf2 signaling pathway. Res. Vet. Sci. 2022, 152, 717–725.
- Cai, D.; Li, X.; Xu, Q.; Li, H.; Liu, R.; Chen, J.; Jiang, X.; Sun, J.; Lai, C.; Bai, W. Cyanidin-3-O-glucoside and protocatechuic acid alleviate heat stress-induced testicular damage. Food Funct. 2023, 14, 2200–2211.
- Mbegbu, E.C.; Ikele, C.G.; Obidike, I.R.; Parrish, J.J. Immunomodulatory potentials of saccharomyces cerevisiae in mitigation of testicular heat stress alterations. J. Reprod. Immunol. 2023, 159, 104057.
- Wang, K.; Li, Z.; Li, Y.; Li, X.; Suo, Y.; Li, C. Impacts of elevated temperature on morphology, oxidative stress levels, and testosterone synthesis in ex vivo cultured porcine testicular tissue. Theriogenology 2023, 212, 181–188.
- Cai, H.; Qin, D.; Peng, S. Responses and coping methods of different testicular cell types to heat stress: Overview and perspectives. Biosci. Rep. 2021, 41, BSR20210443.
- Wang, C.; He, C.; Gao, Y.; Wang, K.; Liang, M. Heat exposure promotes apoptosis and pyroptosis in Sertoli cells. Biocell 2023, 47, 155–164.
- Deng, C.C.; Zhang, J.P.; Huo, Y.N.; Xue, H.Y.; Wang, W.; Zhang, J.J.; Wang, X.Z. Melatonin alleviates the heat stress-induced impairment of Sertoli cells by reprogramming glucose metabolism. J. Pineal Res. 2022, 73, e12819.
- Xue, H.; Huo, Y.; Hu, Y.; Zhang, J.; Deng, C.; Zhang, J.; Wang, X. The role of ALOX15B in heat stress-induced apoptosis of porcine Sertoli cells. Theriogenology 2022, 185, 6–15.
- Hu, Y.; Hu, H.; Yin, L.; Wang, L.; Luo, K.; Luo, N. Arachidonic acid impairs the function of the blood-testis barrier via triggering mitochondrial complex-ROS-P38 MAPK axis in hyperthermal Sertoli cells. Ecotoxicol. Environ. Saf. 2023, 252, 114598.
- He, C.; Sun, J.; Yang, D.; He, W.; Wang, J.; Qin, D.; Zhang, H.; Cai, H.; Liu, Y.; Li, N.; et al. Nrf2 activation mediates the protection of mouse Sertoli Cells damage under acute heat stress conditions. Theriogenology 2022, 177, 183–194.
- Monageng, E.; Offor, U.; Takalani, N.B.; Mohlala, K.; Opuwari, C.S. A Review on the Impact of Oxidative Stress and Medicinal Plants on Leydig Cells. Antioxidants 2023, 12, 1559.
- Xiong, Y.; Li, J.; He, S. Zinc Protects against Heat Stress–Induced Apoptosis via the Inhibition of Endoplasmic Reticulum Stress in TM3 Leydig Cells. Biol. Trace Elem. Res. 2022, 200, 728–739.
- Kawano, K.; Sakaguchi, K.; Madalitso, C.; Ninpetch, N.; Kobayashi, S.; Furukawa, E.; Yanagawa, Y.; Katagiri, S. Effect of heat exposure on the growth and developmental competence of bovine oocytes derived from early antral follicles. Sci. Rep. 2022, 12, 8857.
- Wang, Y.R.; Chen, K.L.; Li, C.M.; Li, L.; Wang, G.L. Heme oxygenase 1 regulates apoptosis induced by heat stress in bovine ovarian granulosa cells via the ERK1/2 pathway. J. Cell. Physiol. 2019, 234, 3961–3972.
- De Rensis, F.; Saleri, R.; Garcia-Ispierto, I.; Scaramuzzi, R.; López-Gatius, F. Effects of heat stress on follicular physiology in dairy cows. Animals 2021, 11, 3406.
- Li, H.; Cong, X.; Sui, J.; Jiang, Z.; Fu, K.; Huan, Y.; Cao, R.; Tian, W.; Feng, Y. Baicalin enhances the thermotolerance of mouse blastocysts by activating the ERK1/2 signaling pathway and preventing mitochondrial dysfunction. Theriogenology 2022, 178, 85–94.
- Sammad, A.; Luo, H.; Hu, L.; Zhu, H.; Wang, Y. Transcriptome reveals granulosa cells coping through redox, inflammatory and metabolic mechanisms under acute heat stress. Cells 2022, 11, 1443.
- Cardone, D.A.; Cáceres, A.R.; Sanhueza, M.A.; Bruna, F.A.; Laconi, M.R. Effects of short-term in vitro heat stress on bovine preantral follicles. Livest. Sci. 2022, 264, 105076.
- Zhang, S.X.; Wang, D.L.; Qi, J.J.; Yang, Y.W.; Sun, H.; Sun, B.X.; Liang, S. Chlorogenic acid ameliorates the heat stress-induced impairment of porcine Sertoli cells by suppressing oxidative stress and apoptosis. Theriogenology 2024, 214, 148–156.
- Bai, L.; Zhang, Y.; Zheng, C.; Xu, S.; He, Y.; Yu, G.; Huang, D.; Huang, Y.; Li, M.; Xu, C. Tanshinone IIA protects mouse testes from heat stress injury by inhibiting apoptosis and TGFβ1/Smad2/Smad3 signaling pathway. Cell Stress Chaperones 2023, 28, 749–759.
- Xiong, Y.; Jin, E.; Yin, Q.; Che, C.; He, S. Boron attenuates heat stress–induced apoptosis by inhibiting endoplasmic reticulum stress in mouse granulosa cells. Biol. Trace Elem. Res. 2021, 199, 611–621.
- Stamperna, K.; Giannoulis, T.; Dovolou, E.; Kalemkeridou, M.; Nanas, I.; Dadouli, K.; Moutou, K.; Mamuris, Z.; Amiridis, G.S. Heat Shock Protein 70 improves in vitro embryo yield and quality from heat stressed bovine oocytes. Animals 2021, 11, 1794.
- Jais, A.; Einwallner, E.; Sharif, O.; Gossens, K.; Lu, T.T.; Soyal, S.M.; Medgyesi, D.; Neureiter, D.; Paier-Pourani, J.; Dalgaard, K.; et al. Heme oxygenase-1 drives metaflammation and insulin resistance in mouse and man. Cell 2014, 158, 25–40.
- Bindu, S.; Pal, C.; Dey, S.; Goyal, M.; Alam, A.; Iqbal, M.S.; Dutta, S.; Sarkar, S.; Kumar, R.; Maity, P.; et al. Translocation of heme oxygenase-1 to mitochondria is a novel cytoprotective mechanism against non-steroidal anti-inflammatory drug-induced mitochondrial oxidative stress, apoptosis, and gastric mucosal injury. J. Biol. Chem. 2011, 286, 39387–39402.
- Kondo, R.; Gleixner, K.V.; Mayerhofer, M.; Vales, A.; Gruze, A.; Samorapoompichit, P.; Greish, K.; Krauth, M.T.; Aichberger, K.J.; Pickl, W.F.; et al. Identification of heat shock protein 32 (Hsp32) as a novel survival factor and therapeutic target in neoplastic mast cells. Blood 2007, 110, 661–669.
- Zenclussen, M.L.; Jensen, F.; Rebelo, S.; El-Mousleh, T.; Casalis, P.A.; Zenclussen, A.C. Heme oxygenase-1 expression in the ovary dictates a proper oocyte ovulation, fertilization, and corpora lutea maintenance. Am. J. Reprod. Immunol. 2012, 67, 376–382.
- Wang, Y.; Yang, C.; Nahla Abdalla Hassan, E.; Li, C.; Yang, F.; Wang, G.; Li, L. HO-1 reduces heat stress-induced apoptosis in bovine granulosa cells by suppressing oxidative stress. Aging 2019, 11, 5535.
- Faheem, M.S.; Dessouki, S.M.; Abdel-Rahman, F.E.; Ghanem, N. Physiological and molecular aspects of heat-treated cultured granulosa cells of Egyptian buffalo (Bubalus bubalis). Anim. Reprod. Sci. 2021, 224, 106665.
- Faheem, M.S.; Ghanem, N.; Gad, A.; Procházka, R.; Dessouki, S.M. Adaptive and Biological Responses of Buffalo Granulosa Cells Exposed to Heat Stress under In Vitro Condition. Animals 2021, 11, 794.
- Guan, J.Y.; Liao, T.T.; Yu, C.L.; Luo, H.Y.; Yang, W.R.; Wang, X.Z. ERK1/2 regulates heat stress-induced lactate production via enhancing the expression of HSP70 in immature boar Sertoli cells. Cell Stress Chaperones 2018, 23, 1193–1204.
- Liang, Y.; Liu, J.; Feng, Z. The regulation of cellular metabolism by tumor suppressor p53. Cell Biosci. 2013, 3, 9.
- Cui, W.; Li, B.; Bai, Y.; Miao, X.; Chen, Q.; Sun, W.; Tan, Y.; Luo, P.; Zhang, C.; Zheng, S.; et al. Potential role for Nrf2 activation in the therapeutic effect of MG132 on diabetic nephropathy in OVE26 diabetic mice. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E87–E99.
- Cullinan, S.B.; Gordan, J.D.; Jin, J.; Harper, J.W.; Diehl, J.A. The Keap1-BTB protein is an adaptor that bridges Nrf2 to a Cul3-based E3 ligase: Oxidative stress sensing by a Cul3-Keap1 ligase. Mol. Cell. Biol. 2004, 24, 8477–8486.
- Pankiv, S.; Clausen, T.H.; Lamark, T.; Brech, A.; Bruun, J.-A.; Outzen, H.; Øvervatn, A.; Bjørkøy, G.; Johansen, T. p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy. J. Biol. Chem. 2007, 282, 24131–24145.
- Lau, A.; Zheng, Y.; Tao, S.; Wang, H.; Whitman, S.A.; White, E.; Zhang, D.D. Arsenic inhibits autophagic flux, activating the Nrf2-Keap1 pathway in a p62-dependent manner. Mol. Cell. Biol. 2013, 33, 2436–2446.
- Lau, A.; Wang, X.-J.; Zhao, F.; Villeneuve, N.F.; Wu, T.; Jiang, T.; Sun, Z.; White, E.; Zhang, D.D. A noncanonical mechanism of Nrf2 activation by autophagy deficiency: Direct interaction between Keap1 and p62. Mol. Cell. Biol. 2010, 30, 3275–3285.
- Copple, I.M.; Lister, A.; Obeng, A.D.; Kitteringham, N.R.; Jenkins, R.E.; Layfield, R.; Foster, B.J.; Goldring, C.E.; Park, B.K. Physical and functional interaction of sequestosome 1 with Keap1 regulates the Keap1-Nrf2 cell defense pathway. J. Biol. Chem. 2010, 285, 16782–16788.
- Li, Z.; Li, Y.; Zhou, X.; Dai, P.; Li, C. Autophagy involved in the activation of the Nrf2-antioxidant system in testes of heat-exposed mice. J. Therm. Biol. 2018, 71, 142–152.
- Rotimi, D.E.; Ojo, O.A.; Olaolu, T.D.; Adeyemi, O.S. Exploring Nrf2 as a therapeutic target in testicular dysfunction. Cell Tissue Res. 2022, 390, 23–33.
- Li, Y.; Cao, Y.; Wang, F.; Pu, S.; Zhang, Y.; Li, C. Tert-butylhydroquinone attenuates scrotal heat-induced damage by regulating Nrf2-antioxidant system in the mouse testis. Gen. Comp. Endocrinol. 2014, 208, 12–20.
- Li, Y.; Cao, Y.; Wang, F.; Li, C. Scrotal heat induced the Nrf2-driven antioxidant response during oxidative stress and apoptosis in the mouse testis. Acta Histochem. 2014, 116, 883–890.
- Li, Y.; Huang, Y.; Piao, Y.; Nagaoka, K.; Watanabe, G.; Taya, K.; Li, C.M. Protective effects of nuclear factor erythroid 2-related factor 2 on whole body heat stress-induced oxidative damage in the mouse testis. Reprod. Biol. Endocrinol. 2013, 11, 23.
- Sui, J.; Feng, Y.; Li, H.; Cao, R.; Tian, W.; Jiang, Z. Baicalin protects mouse testis from injury induced by heat stress. J. Therm. Biol. 2019, 82, 63–69.
- Feng, J.; He, Y.; Shen, Y.; Zhang, G.; Ma, S.; Zhao, X.; Zhang, Y. Protective effects of nuclear factor erythroid 2-related factor on oxidative stress and apoptosis in the testis of mice before adulthood. Theriogenology 2020, 148, 112–121.
- Murata, H.; Kunii, H.; Kusama, K.; Sakurai, T.; Bai, H.; Kawahara, M.; Takahashi, M. Heat stress induces oxidative stress and activates the KEAP1-NFE2L2-ARE pathway in bovine endometrial epithelial cells. Biol. Reprod. 2021, 105, 1114–1125.
- Li, J.; Zhao, W.; Zhu, J.; Ju, H.; Liang, M.; Wang, S.; Chen, S.; Ferreira-Dias, G.; Liu, Z. Antioxidants and Oxidants in Boar Spermatozoa and Their Surrounding Environment Are Associated with AMPK Activation during Liquid Storage. Vet. Sci. 2023, 10, 214.
- Froment, P.; Plotton, I.; Giulivi, C.; Fabre, S.; Khoueiry, R.; Mourad, N.I.; Horman, S.; Rame, C.; Rouillon, C.; Grandhaye, J.; et al. At the crossroads of fertility and metabolism: The importance of AMPK-dependent signaling in female infertility associated with hyperandrogenism. Hum. Reprod. 2022, 37, 1207–1228.
- Yang, W.; Wang, L.; Wang, F.; Yuan, S. Roles of AMP-activated protein kinase (AMPK) in mammalian reproduction. Front. Cell Dev. Biol. 2020, 8, 593005.
- Meroni, S.B.; Galardo, M.N.; Rindone, G.; Gorga, A.; Riera, M.F.; Cigorraga, S.B. Molecular mechanisms and signaling pathways involved in sertoli cell proliferation. Front. Endocrinol. 2019, 10, 224.
- Ni, F.D.; Hao, S.L.; Yang, W.X. Multiple signaling pathways in Sertoli cells: Recent findings in spermatogenesis. Cell Death Dis. 2019, 10, 541.
- Yang, W.R.; Liao, T.T.; Bao, Z.Q.; Zhou, C.Q.; Luo, H.Y.; Lu, C.; Pan, M.H.; Wang, X.Z. Role of AMPK in the expression of tight junction proteins in heat-treated porcine Sertoli cells. Theriogenology 2018, 121, 42–52.
- Bertoldo, M.J.; Guibert, E.; Faure, M.; Guillou, F.; Ramé, C.; Nadal-Desbarats, L.; Foretz, M.; Viollet, B.; Dupont, J.; Froment, P. Specific deletion of AMP-activated protein kinase (α1AMPK) in mouse Sertoli cells modifies germ cell quality. Mol. Cell. Endocrinol. 2016, 423, 96–112.
- Xiao, L.; Fang, Z.; Wang, Q.; Sheng, X.; Qi, X.; Xing, K.; Guo, Y.; Ni, H.; Wang, X.; Zhang, Y. Curcumin Ameliorates Age-Induced Tight Junction Impaired in Porcine Sertoli Cells by Inactivating the NLRP3 Inflammasome through the AMPK/SIRT3/SOD2/mtROS Signaling Pathway. Oxid. Med. Cell. Longev. 2023, 2023, 1708251.