Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Kleber Del-Claro and Version 2 by Rita Xu.
The net outcomes of mutualisms are mediated by the trade-offs between the costs and benefits provided by both partners. This trade-off has important implications for understanding the evolution of extrafloral nectaries (EFNs), an adaptation that has repeatedly evolved throughout the flowering plant clade.
- protective mutualism
- extrafloral nectar
- ant–plant interaction
1. Ant Protection Mutualisms Mediated by Plant-Based Resources
The nature of interspecific interactions, whether positive or negative, depends on the balance sheet of the contributions of each species to these interactions. In protective mutualisms, ants are attracted to foraging on plants because the plants provide food rewards (Figure 1A,B), domiciles, or both [1][2][3][4][5][6][7][8][1,2,3,4,5,6,7,8]. In turn, when acting as mutualists (Figure 1C–G), ants harass, bite, eat, chase off, or otherwise dissuade herbivores from eating the plants [3][9][10][11][3,9,10,11] (Figure 1H–J). The plants provide a reward to benefit the ants [10][12][13][14][15][16][17][10,12,13,14,15,16,17], and from the plant’s balance sheet, the cost of the food award or domicile may be outweighed by the benefit of the reduced tissue loss due to reduced herbivory [2][18][2,18]. Meta-analyses reveal that, in general, ants visiting plants reduce herbivory and increase seed production, demonstrating a positive balance sheet [19][20][19,20]. However, not all ant species are equally effective guards, and some are cheaters, providing no protection at all [21][22][21,22].

Figure 1. Extrafloral nectar drops in (A) Stryphnodendron adstringens (Fabaceae) and (B) Qualea multiflora (Vochysiaceae). (C,D) Ectatomma tuberculatum collecting extrafloral nectar. (E) Ectatomma sp. and (F) Neoponera villosa ants collecting nectar from the extrafloral nectaries on the inflorescence of Banisteriopsis malifolia (Malpighiaceae). (G) Camponotus leydigi collecting nectar from the extrafloral nectaries on the inflorescence of Bionia coriacea (Fabaceae). (H) Ectatomma tuberculatum and (I) Camponotus crassus foraging on the flowers of Q. multiflora. (J) Ectatomma tuberculatum foraging on the flowers of Senna rugosa (Fabaceae). Photos: E.S. Calixto, D. Lange, H. M. Torezan-Silingardi, K. Del-Claro.
In protective mutualisms, there is increasing evidence that, in some cases, ants’ interference in the pollination process can be a negative entry or liability on a plant’s balance sheet [21][23][24][25][26][27][28][29][21,23,24,25,26,27,28,29]. Ants, otherwise attracted to plants because of their extrafloral nectar, can rob floral nectar, damage floral parts, reduce pollen viability, and chase away potentially effective pollinators.
In their study experimentally testing the flower distraction hypothesis, Wagner and Kay [30][32] refer to a single study showing that ants influence floral visitors, in this case in a plant species without EFNs. The number of studies on ant–pollinator conflict has increased in the past few years [23][24][25][26][29][31][32][23,24,25,26,29,30,33], although the outcomes of this relationship in terms of plant fitness remain unresolved. TheOur specific goals are to (i) review the known negative effects that ants have on pollination and consequently on plant fitness; (ii) describe the factors that might reduce the impact of ants on the pollination process; and (iii) suggest the existence of “optimal ant partners” (and consequently non-optimal ant partners) in the protection mutualism system, similar to the role of effective pollinators in pollination interactions [33][34]. Importantly, rwesearchers suggest that optimality of the ant partner not only depends on its contribution to plant fitness via protection but is also balanced against its potential negative effect on plant fitness via pollinator dissuasion. (iv) Finally, researcherswe discuss the kinds of plants, specifically in terms of their architectures, phenologies, and floral morphologies, which are most likely to be prone to the negative effects of ants on pollination processes. Overall, ants are attracted by EFNs, suggesting the existence of optimal patterns that have the highest positive effects on plant protection and the lowest negative effects on pollination processes (Figure 2).
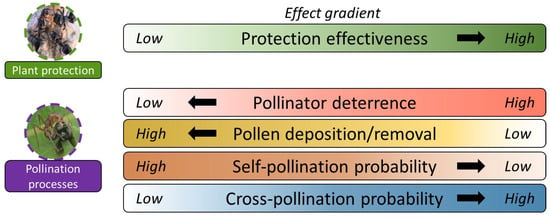
Figure 2. Trade-off between plant protection and disruption of pollination processes. Hypothetical framework for understanding the potential trade-off between plant protection against herbivore attack and negative effects on pollination processes by mutualistic ants attracted by extrafloral nectaries. Ants protect plants against herbivore attacks (positive effect), but due to their different levels of protection effectiveness and other traits (e.g., size, pheromones), they may impair the pollination process, ultimately resulting in a decrease in plant fitness (negative effect). An ant species that has the highest positive effects in protection effectiveness and the lowest negative effects on pollination processes is therefore an optimal mutualist partner (black arrows show the direction of the effects of optimal ant partners on plant protection and pollination processes).
2. Negative Effects of Ants on Pollinators
Although ants can protect reproductive parts [34][35][36][37][35,36,37,38], their presence on flowers may generate negative effects on pollination [38][31], such as when ants repel or prey on potential pollinators, negatively impacting the reproductive success of the plant [23][24][25][26][27][28][23,24,25,26,27,28]. For instance, in the Neotropics, Ectatomma tuberculatum ants occupy certain parts of the plant (Figure 1), such as the EFNs (13 ± 9 s, mean ± SD) and reproductive parts (23 ± 11 s), more often than other ant species, such as Camponotus crassus (EFN: 1 ± 1 s, flower: 2 ± 1 s) [25]. When close to the flowers, E. tuberculatum will deter or even attack pollinators [25]. In general, few studies have evaluated the direct and indirect impacts that aggressive ants like Ectatomma have on pollinators or plant fitness, especially for EFN-bearing plants [24][25][26][27][29][39][40][41][42][24,25,26,27,29,39,40,41,42]. ResWearchers believe that the identity of the ants and pollinators can be very important to this trade-off [25][27][43][25,27,43]. Ant identity can be understood as a combination of shape, size, aggressiveness, and odor, and pollinator identity can be understood as a combination of size, agility, and likelihood of visiting or not visiting a flower occupied by ants [44]. Therefore, the identity of the ants, as well as that of the floral visitors, can impact the fitness of plants in different ways.
Studies have shown that very aggressive ants can deter or even prey on pollinators, significantly changing pollinator behavior and/or reducing fruit and seed set [26][27][26,27]. Ness [27] observed that the visitation rates and duration of visits to the flowers of Ferocactus wislizeni (Cactaceae) by pollinators differed according to the level of aggressiveness of the ants attending the host plants. Flowers occupied by the aggressive ant Solenopsis xyloni were visited less frequently and for less time, resulting in fruits with significantly smaller and lighter seeds compared to those of plants with more docile ants. These results show that very aggressive ants can restrict or decrease the reproduction of plants via the disruption of pollination processes in certain environments; however, see [40].
Some pollinators are able to identify ant body shape and avoid visiting flowers occupied by ants [24][25][45][24,25,45]. Villamil et al. [26] found that in Turnera velutina (Passifloraceae), the presence of ant corpses, experimentally placed in flowers, reduced the duration of visits by pollinators, but these results were contingent on the ant species. The greatest negative effects were caused by the most aggressive ants. In addition, Calixto et al. (in review) studied three species of ants of different sizes and levels of aggressiveness in Qualea multiflora (Vochysiaceae) in the Brazilian savanna. They showed that the presence of dead ants in the flowers, again experimentally placed there, decreased plant fitness by preventing the visitation of potential pollinators. They found that the treatment with the largest and most aggressive ants was the treatment most avoided by pollinators. In experiments using plastic ants, Assunção et al. [25] showed that ants were recognized as a danger to pollinators, Heteropterys pteropetala (Malpighiaceae) with plastic ants produced fewer fruits than flowers with plastic circles or control flowers. In a similar study, Nogueira et al. [24] showed that pollinators hesitated to visit flowers with artificial ants, negatively affecting pollination, but did not hesitate to visit flowers with plastic circles, suggesting that they recognized the specific morphology of the ants. Also, they showed that pollinators spent 2.5 times more time per flower on the ant-free branches, and the number of fruits produced per flower was 2 times lower in the group with plastic ants.
In addition to size and aggressiveness, the number of ants foraging on plants and the pheromones they release are indicators used by pollinators. Large numbers of ant individuals (workers and/or soldiers) might be seen from far distances, dissuading pollinators from visiting flowers [44]. For instance, using field observations and experimental manipulations of ant density in the branches of Banisteriopsis malifolia (Malpighiaceae), Barônio and Del-Claro [44] demonstrated a significant decrease in the number of visits and fruit set as the density of ants increased. Ants release trail and territorial pheromones produced by different glands, such as the metapleural glands, to communicate or dominate territories [46][47][48][46,47,48]. Pollinators, in turn, can learn to use these scent marks as informational signs or associate them with the risk of attack or danger, avoiding visiting plants with ants [49][50][51][49,50,51]. Ballantyne and Willmer [50] demonstrated in laboratory tests that bee drones learn to use ant scent marks as informational signals during foraging on artificial flowers. In another study, Cembrowski et al. [49] showed that Bombus impatiens bees collect less pollen from artificial flowers that ants have crawled on compared to controls, suggesting that some bees use ant scent marks to avoid conflict with ants when visiting flowers.
Finally, the pollinator responses also vary depending on the pollinator identity. Some pollinators are more likely to visit flowers containing ants than other pollinators. For instance, in a study conducted by Barônio and Del-Claro [44], the presence of ants around the flowers of Malpighiaceae plants (e.g., Figure 1E,F) reduced the visitation rates of smaller non-Centridini bees but not larger (usually >1 cm) Centridini bees. Studying the interaction between the weaver ant Oecophylla smaragdina and the tropical shrub Melastoma malabathricum, Gonzálvez et al. [52] observed that smaller bees tended to visit ant-free plants, while large bees concentrated their visits on plants occupied by ants. Other studies have also shown this pattern, in which smaller bees tend to approach more carefully flowers containing ants [25][44][49][53][25,44,49,53], reinforcing the evidence that the identity of the floral visitors in addition to the identity of the ants is also a factor determining the trade-off between protection and deterrence.
3. Positive Effects of Pollinator Repellence by Ants
Although there seems to be a clear negative effect of the presence of ants when foraging on flowers on pollinator behavior, a few studies have shown positive outcomes [23][32][51][54][23,33,51,54]. Decreasing pollinator visitation to flowers in response to the presence of ants may lead to decreased self-pollination or may increase the likelihood of cross-pollination, as pollinators are forced to constantly change flowers within plants and/or to move to other plants due to the presence of ants [23][32][51][54][23,33,51,54]. In this context, there is a decrease in the foraging efficiency of pollinators on the same plant individual but an increase in movement between individual plants, bringing positive effects for plant reproductive success. Villamil et al. [29] provided evidence of the deterrence of floral visitors by ants in an ant–plant interaction mediated by EFNs. Studying the shrub Turnera velutina (Passifloraceae), they showed that the more aggressive the ant, the more likely it is to deter floral visitors; however, they also showed that this deterrence reduced three-fold self-pollination and increased cross-pollination two-fold (also see [51]). ResWearchers are just beginning to understand what the net outcomes of this trade-off between protection from herbivore attack and pollinator deterrence are, as well as which biotic factors may affect the outcomes of these interactions. By addressing these questions, researcherswe can elucidate the costs and benefits, and therefore the net outcomes, provided by ant mutualists to plants bearing EFNs.