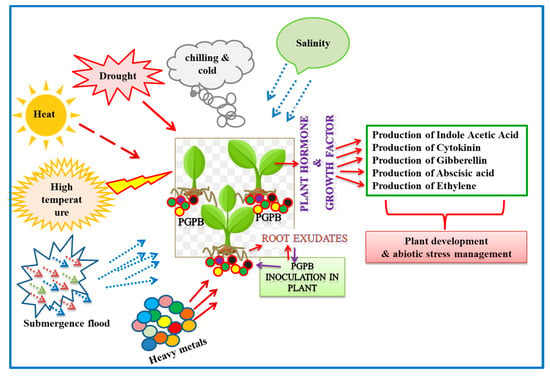
2. Dynamics of Plant-Microbe Interactions in Extreme Habitats
In the extreme environment, the microbes thrive in the plant vicinity and form symbiotic associations with their plant counterparts. Exhibiting diversity, the microbes are ubiquitous and perform diverse functions, including nutritional uptake, conferring tolerance to environmental stress, and promoting plant growth and development. The existence of microbes in extreme niches raises curiosity to explore the dynamics of symbiotic association, their environmental impact, and biotechnological utilities
[33][35] (
Figure 1).
Figure 1. Diagrammatic representation of plant growth-promoting bacteria (endophytic, rhizospheric, and epiphytic) and functional attributes in impacting plant growth, development, and abiotic stress (viz. heat, drought, salinity, chilling, and flood) management.
In extreme habitats, the microbes have evolved distinct genetic and physiological mechanisms to survive and adapt to challenging environmental conditions
[8][34][8,36]. To get a better understanding, microbes have been isolated and characterized from extreme environments by both culture-dependent and culture-independent techniques
[7][35][36][7,37,38] and meta-omics approaches
[37][34].
3. Diversity and Distribution of Plant-Associated Extremophiles
3.1. Epiphytic Microbiomes
The above-ground microbial communities (bacteria, fungi, and yeast) in plant association are referred to as the phyllospheric microbes
[38][57]. The aerial zone inhabited by the microbes is known as phyllosphere and the microbes are designated as epiphytes. Mostly, the bacterial communities densely inhabit the leaf surface (epiphytes), however, leaf surface colonization by epiphytes represents a challenging zone for colonization
[39][40][58,59]. The microbial communities colonizing leaves include different genera of bacteria, algae, filamentous algae, and protozoans. The bacterial species are the most abundant in the phyllosphere and are found in approximately 10
6 to 10
7 cells/cm
2 of the leaf
[41][60]. The epiphytic bacterial communities exhibit size differences among plant species caused by the physical and nutritional condition of the phyllosphere
[40][59]. Moreover, epiphytic microbes have distinct characteristics and show tolerance to high temperatures (40–55 °C) and ultraviolet radiation, being constantly exposed to adverse environmental conditions. Microbes from different phyllosphere zones of crops were reported:
Agrobacterium,
Methylobacterium,
Pantoea,
Bacillus,
Enterobacter, and
Pseudomonas comprising key microbial species in moderate and extreme environmental conditions
[40][59]. Studies have suggested that leaf-colonizing bacteria promote plant growth and confer tolerance to environmental stresses
[42][61].
3.2. Endophytic Microbiomes
Endophytes constitute bacterial or fungal microorganisms, inhabiting inter/intracellular spaces within plant tissues, and present in almost all plant species. Since plants restrict the growth of endophytes, endophytes evolve mechanisms for adaptation to the environment
[43][44][69,70], including the production of metabolites for plant growth and development
[9][10][11][9,10,11]. Endophytes have been isolated from different plant parts namely meristem, leaves
[45][71], seeds
[46][72], roots
[36][38], and stem, among other tissues. However, studies have suggested that isolation of some endophytes is difficult, and different methods have been employed, namely plant tissue culture on suitable media
[46][72] and endophyte isolation using surface sterilization of ground tissue extract
[47][73]. The fungal endophytes from plants and algae are classified in
Ascomycetes, while very few reports on
Basidiomycetes are available
[48][74]. Some of the microorganisms, namely
Penicillium glandicola,
Acremonium terricola, and
Phoma tropica, were classified as fungal endophytes
[49][75]. The endophytic microbial communities comprised of
Achromobacter,
Azoarcus,
Burkholderia,
Enterobacter,
Gluconoacetobacter,
Herbaspirillum, etc. were isolated from different plant species
[17][18][17,18]. Plant endophyte associations have important ecological and biotechnological attributes
[50][51][52][76,77,78].
3.3. Rhizospheric Microbiomes
The rhizospheric microbial communities present in extreme environments, i.e., drought, salinity, and acidity/alkalinity have developed adaptive mechanisms for survival and are characterized
[26][53][26,79]. The microbial communities belonging to archaeal phyla
Euryarchaeota and
Crenarchaeota and bacterial phyla
Actinobacteria,
Bacteroidetes,
Proteobacteria, and
Firmicutes) were predominantly present in plant rhizosphere, in extreme habitats
[54][80]. The
Proteobacteria consist of
α/
β/
γ/
δ-
Proteobacteria, found in close association with most of the crop plants. The agriculturally significant species comprise of
α-
Proteobacteria which requires low nutrients for growth and induces nitrogen fixation in plant symbiosis. The
β-
Proteobacteria have a high metabolic rate and the
γ-
Proteobacteria define the largest class, including
Azotobacter and
Pseudomonas. The genus
Azospirillum is closely associated with crop plants namely
Amaranthus, sorghum, sugarcane, maize, and ryegrass, demonstrating symbiotic nitrogen fixation
[55][81].
4. Plant-Microbe Interactions in Extreme Ecological Habitats
4.1. Acidic Environment
Acidophiles comprise microbes that thrive in highly acidic environments, as low as pH < 3. Acidophiles are found in diverse ecological niches, including hydrothermal regions, volcanic areas, deep-sea vents, and the stomachs of animals
[56][57][91,92]. High acidic conditions are found to adversely affect plant growth, leading to changes in the availability of nutrients and soil pH. The plant counters high acidic conditions by maintaining its internal pH. Diverse rhizospheric microbes inhabit acidic environments: acid-tolerant PGPB including
Acidithiobacillus,
Flavobacterium,
Lysinibacillus,
Pseudomonas, and
Methylobacterium were isolated and characterized
[58][93]. The best-characterized acidophiles are classified in Archea and bacterial domains
[59][94]. The microbial communities associated with the crop plants are found to be essential for soil health and crop productivity
[60][61][95,96] and siderophore production alleviates abiotic stress conditions.
4.2. Alkaline Environment
The microbes from alkaline environments can tolerate high pH (>9) and are known as alkaliphiles. These extremophiles possess genetic and physiological mechanisms to survive in harsh conditions. The rhizospheric zone of the plant colonizing alkaline environments such as
Smallanthus sonchifolius,
Dichanthium annulatum, and
Chrysanthemum morifolium comprises diverse microbes from methane and hydrogen-rich environments
[54][80]. In these alkaline conditions, microbes maintain cytoplasmic pH through protein and enzyme activity. Moreover, the alkaliphilic bacteria have the adaptive mechanism to tolerate a diverse range of pH 6–10, with pH 7–8 being optimum for growth
[62][100].
4.3. Drought Condition
In adverse environmental conditions, plant adaptation and survival are promoted by AM fungi and PGP microbes
[63][102]. In this regard, the drought-tolerant microbes have evolved to adapt/survive in water-deficit conditions and protect host plants by facilitating nutrient uptake and plant growth. The microbes colonize the rhizospheric zone and employ multiple direct or indirect mechanisms including the production of ACC deaminase, phytohormones (abscisic acid, cytokinins, and IAA), bacterial exopolysaccharides, and induced systemic tolerance
[64][103].
The PGPR produces phytohormones that promote plant growth in stress conditions, for example, IAA governs the differentiation of vascular tissues, and cell division, and promotes the growth of shoots under drought stress
[65][110]. On the other hand, the ABA hormone mitigates drought stress by increasing the transcription of drought-linked genes
[66][111]. ACC deaminase (from bacteria) hydrolyzes ACC into alphaketobutyrate and ammonia
[67][112]. The PGP and drought-tolerant bacteria enhance water potential, and biomass, thereby minimizing water loss in drought conditions
[68][113]. The rhizospheric microbes produce exopolysaccharides and alleviate drought stress in plants
[69][114]. In
Lycopersicum esculentum, phytohormone strigolactone production was increased on
Rhizophagus irregularis colonization and exposure to drought, showing a signaling mechanism of the phytohormone in increasing drought tolerance
[70][115]. The drought-resistant rhizobacteria modulate phytohormones and confer drought tolerance- PGPRs (
Acinetobacter,
Bacillus thuringiensis,
Azospirillum, etc.) synthesize IAA that alters root architecture by augmenting root surface area and root tips, nutrient acquisition, and aid plant in overcoming drought
[71][72][116,117].
4.4. High Temperature
High-temperature conditions adversely affect plant growth by altering membrane permeability, seed germination, and rate of photosynthesis
[73][123]. The rhizospheric microbes of plants inhabiting extremely high temperatures,
Triticum aestivum,
Cupressus dupreziana, and
Sporobolus indicus, promote plant growth in hot conditions. PGPB increases plant growth through several mechanisms, comprising nitrogen fixation, solubilization of P, and Zn, phytohormone production, HCN, and siderophore production
[74][124]. Due to the ability of PGPB in plant growth, many bacterial genera namely
Arthrobacter,
Streptomyces,
Pseudomonas, and
Staphylococcus sp. are used as bioformulations for plant growth in high-temperature conditions. In high temperatures, the metabolism and physiology of extremophiles are adversely affected, and the microbial enzymes promote high-temperature acclimatization and protection of cell structure and integrity via increased expression of heat-tolerant proteins
[8]. Several fungal species have been isolated from hot habitats and comprise
Talaromyces thermophilus,
T. byssochlamydoides,
Malbranchea cinnamomea,
Aspergillus terreus,
Myceliophthora fergusii,
Thermomyces lanuginosus [75][125],
Myceliophthora thermophila [76][126],
Scytalidium thermophilum [77][127], and others
[78][128]. The fungal communities protect the plant by performing several functions, which include P, potassium (K), and Zn solubilization, phytohormone production, and siderophore production for plant adaptation and survival
[60][61][79][84,95,96].
4.5. Low Temperature
The microbes inhabiting extremely cold temperatures are designated as psychrotrophic microbes and have prospects in medicine, agriculture, and industries. Microbes from cold habitats are universally present, found in mountain caps, glaciers, frozen lakes, and snow, and in association with plants growing in cold habitats. Diverse microbes were isolated by culture-dependent and culture-independent techniques and classified as
viz.,
Euryarchaeota,
Ascomycota,
Basidiomycota,
Chlamydiae,
Cyanobacteria,
Actinobacteria,
Chloroflexi,
Bacteroidetes, etc.
[80][134]. The microbes inhabiting cold climates define importance in the ecological perspective since a considerable portion of aquatic and terrestrial ecosystems are influenced by cold temperatures. Moreover, cold regions extremophiles have been reported from Antarctica and extreme cold regions of the world. The extremophiles show extreme diversity and novel psychrophilic microbes comprise
Oleispira antarctica [81][135],
Flavobacterium frigidarium [82][136],
Octadecabacter arcticus [83][137],
Sphingomonas glacialis [84][138],
Halobacterium lacusprofundi [85][139], and
Cellulophaga algicola [86][140].
4.6. Saline Condition
Globally, most of the land in agriculture is threatened by the presence of saline conditions that result in poor microbial functions, due to osmotic stress and ion-induced toxicity
[64][103] detrimental to plant growth. Moreover, soil salinity adversely affects the plants including seed germination, uptake of nutrients and water, crop productivity, and ecological balance
[87][142]. Several studies demonstrated the beneficial effect of PGP and endophytic microbes in mitigating the negative effect of salinity in soil
[88][143]. In a key example, the PGPB
Pseudomonas stutzeri, when inoculated in salt-sensitive and tolerant plants, reduced the adverse effects
[89][144]. Sometimes, inoculation of salt-tolerant bacteria together with AM fungi considerably improves the plant tolerance to salinity stress
[90][145]. Salinity-tolerant microbes adopt several direct and indirect mechanisms to counter salinity stress, and these include the production of phytohormones, mobilization of nutrients, nitrogen fixation, and siderophore production
[91][146]. These microbial mechanisms contribute to root length increase, number of roots, and surface area by uptake of nutrients
[92][147].
4.7. Presence of Heavy Metals
The presence of heavy metals in agrosystems has intensified across the globe and the high concentration of heavy metals is toxic and adversely impacts plant growth and functions. The decrease in crop yield affects human health and food demands, cadmium (Cd) and lead (Pb) are the major toxic heavy metals affecting
O. sativa, a staple food crop. In addition, the accumulation of heavy metals in crops can cause serious health damage.
In recent times, heavy metal stress alleviation employing microbes is gaining importance, and key examples include
Pseudoalteromonas sp.,
Bacillus,
Salmonella sp.
[93][94][95][152,153,154]. In heavy metal-stress plants, rhizobacteria produce IAA and elevate plant growth in polluted soil via macro and micronutrient uptake and conferring plant tolerance to heavy metals
[72][117]. An interesting example is
Deinococcus radiodurans, an extremophile bacterium that occurs in soil
[96][155]. The bacteria have a high concentration of Mn
2+-metabolite complexes that can scavenge ROS
[97][156]. The inoculation of rice plants with the bacteria releases antioxidants that improve plant tolerance to Pb and Cd stress.
4.8. Flooding Condition
In the fluctuating climate scenario, the increase in flood conditions poses havoc for land plants and causes flooding stress. The flooding stress adversely impacts the plants and causes metabolic and physiological changes and alters the plant-associated microbiome. The nature-based solutions to tackle flooding stress utilize living organisms to minimize the effects of climatic fluctuations
[98][99][158,159].
5. Biotechnological Applications of Plant Microbiome
5.1. Plant Growth Promotion
5.1.1. Production of Phytohormones
Phytohormones play an integral role in affecting plant growth dynamics via multiple physiological and biochemical changes in the plant life cycle
[100][101][184,185]. In the mitigation of biotic and abiotic stresses, PGPB found in the rhizospheric zone secretes many phytohormones and modulates the concentration of specific growth hormones in the plant
[102][182]. In the rhizospheric zone, different rhizosphere colonizing bacteria were shown to produce phytohormones to enhance plant growth
[103][104][186,187]. Phytohormones are chemical messengers that in small amounts regulate cellular activities, key examples include abscisic acid, cytokinin, auxins, brassinosteroids, and jasmonates, etc. and some are key targets for plant metabolic engineering for conferring abiotic stress tolerance
[105][106][188,189].
Cucumis sativus root secretes vanillic acid and p-coumaric acid which demonstrate differential effects on the soil microbiome.
Rhizosphere colonizing or endophytic ACC deaminase-producing bacteria alters plant ethylene levels
[107][194]. Abscisic acid (ABA) greatly assists plants in countering environmental stresses and is actively involved in various defense mechanisms. Shahzad et al.
[108][195] investigated the favorable impact of
Bacillus amyloliquefaciens inoculation in rice on plant growth attributes in salinity conditions.
5.1.2. Biological Nitrogen Fixation
Nitrogen is a very important element for plant growth since it plays a major role in amino acid synthesis, the key building blocks of proteins, and is a major component of chlorophyll, an important pigment for photosynthesis. It is also found in other important biomolecules, such as nitrogen bases including nucleotides and nucleosides (ATP, GTP, CTP, and TTP, etc.) and nucleic acids. Prokaryotic organisms possess a widespread ability to fix atmospheric nitrogen
[109][202]. PGPB positively impacts plant growth via- direct mechanisms, including multiple processes such as nitrogen fixation, phosphate solubilization, production of siderophore, ammonia, and phytohormones, etc. while the indirect mechanisms comprise antibiotic production, ACC deaminase activity, induced systemic resistance (ISR) among others
[110][203].
5.1.3. Mineral Solubilization
Plant well-being is greatly influenced by mineral nutrients and during abiotic stress, plants are unable to absorb the minerals and micronutrients severely hampering plant growth and leading to plant disease. The above damages are corrected naturally by microbes that can convert complex forms to simple forms that are easily absorbed by the plants (e.g., siderophores). Siderophores are produced by bacteria, fungi, and plants to facilitate the uptake of iron
[111][112][207,208] and function as iron chelators (bind iron present in the rhizosphere).
The poor availability of inorganic phosphate (orthophosphate) in soil hampers crop production
[113][209]. The phosphate solubilizing bacteria converts insoluble inorganic phosphate
[114][210] to soluble forms and improves phosphorous availability for the plant. Joe and coworkers
[115][211] isolated
Acinetobacter sp. and
Bacillus sp. from
Phyllanthus amarus which showed phosphate solubilization and salt tolerance and increased plant growth compared to non-inoculated plants.
5.1.4. Biocontrol Function
Biocontrol agents secrete biochemical and other substances and inhibit harmful pathogenic bacteria without damaging plants and soil. The plants/crops are disease-affected by reducing crop yields, contamination of food grains, and declining production quality. Multiple PGPB synthesizes salicylic acid that signals systemic acquired resistance (SAR) while PGPB may start to induce systemic resistance (ISR), enhancing plant defense against plant pathogens
[116][180]. PGPBs are key players in disease management, maintain ecological subsistence, and reduce the deleterious effects of chemical fertilizers
[117][118][212,213]. The representative examples include
Arthrobacter,
Enterobacter,
Pseudomonas,
Rhizobium, and
Frankia spp.
5.2. Mitigation of Multiple Abiotic Stress
Recent agricultural trials have scientifically validated that PGPRs not only reduce environmental stresses but also increase the production of a variety of crop plants, such as soybeans, mint, rice, barley, and maize
[91][119][120][121][146,214,215,216]. Hormones primarily control the prioritization of signals carried out by protein switches such as kinases, transcription factors (TFs), and G-proteins, according to molecular research (gene expression profiling). Usually, plants focus their physiological resources on abiotic stress adaptation, which renders them prone to biotic stressors such as herbivory and disease
[122][217].
The hormone that is primarily involved in the abiotic stress response is ABA. Plants respond to abiotic stress through defense mechanisms activated by ethylene, salicylic acid, or jasmonate
[123][218].
Microbe-mediated development of abiotic stress responses is often referred to as induced systemic tolerance (IST). Over the past few decades, there has been a lot of research conducted regarding the function that microbes play in helping plants cope with abiotic challenges
[124][219]. Plants experience less abiotic stress thanks to the possible inherent metabolic and genetic capacities of microbes
[102][116][180,182].
T. harzianum function in the rice genotype decreases stress through the overexpression of physiological genes, specifically those encoding aquaporin, dehydrin, and malondialdehyde
[125][175]. The rhizosphere makes up a soil microclimate around the root zone, where the average number of microbes is significantly higher than in the bulk soil. Therefore, it is evident that a variety of nutrients, minerals, and metabolites found in plant roots may play a significant role in drawing microbes to gather and form partnerships with plants.
5.2.1. Heat Stress
The microbes belonging to different genera including
Azospirillum,
Achromobacter,
Variovorax,
Enterobacter,
Bacillus,
Azotobacter,
Klebsiella,
Aeromonas, and
Pseudomonas promote plant growth under heat conditions
[126][220].
T. aestivum inoculated with
Azospirillum brasilence, and
Bacillus amyloliquefaciens, under heat stress resulted in reduced regeneration of ROS (reactive oxygen species), pre-activation of heat shock transcription factors, and changes in metabolome
[127][221].
5.2.2. Cold Stress
The freezing (cold) damage is one of the main causes of crop loss
[128][129][222,223]. It lowers crop production and productivity by slowing down plant growth and development
[130][131][224,225]. PGPRs are beneficial to many plants as they increase their resistance to various stressors, such as low temperatures. Su et al.
[132][226] showed that
Burkholderia phytofirmans PsJN decreased the effect of freezing temperatures on
A. thaliana photosynthesis.
5.2.3. Drought Stress
According to Mittler et al.
[133][227] and Cramer et al.
[2], agricultural loss occurs in different crops due to abiotic stress namely water deficit (drought) conditions, and affects 64% of the global land area, respectively. The potential of microbial interactions with the plants has, therefore, multifaceted functions, one of them is adaptation under drought stress. The root fungal endophyte
Piriformospora indica induces drought tolerance in Chinese cabbage by increasing the levels of antioxidants and improving many physiological parameters
[134][228].
5.2.4. Salinity Stress
T. harzianum application to increase the oil content in NaCl-affected Indian mustard (
Brassica juncea), improved nutrient uptake, and improved the accumulation of antioxidants and osmolytes while decreasing NaCl uptake
[135][229]. Concurrent with these results, it was shown that plants treated with
Trichoderma produced higher levels of monodehydroascorbate reductase. Additionally, research on mutants has verified that
Trichoderma produces ACC-deaminase, which reduces the effects of salt stress
[136][230].
Pseudomonas sp. and
Acinetobacter sp. have been shown to increase the production of IAA and ACC deaminase in salt-affected soil in oats and barley
[137][231].
B. phytofirmans strain PsJN mitigates drought stress in maize
[138][232], wheat
[139][233], and salt stress in
Arabidopsis [140][234]. Salt tolerance in rice variety improved in germination under salinity stress via
Pseudomonas sp. inoculation.