Pyrrolizidine alkaloids (PAs) are naturally occurring secondary metabolites of plants. More than 660 types of PAs have been identified from an estimated 6000 plants, and approximately 120 of these PAs are hepatotoxic. As a result of PAs being found in spices, herbal teas, honey, and milk, PAs are considered contaminants in foods, posing a potential risk to human health.
- pyrrolizidine alkaloids
- hepatotoxicity
- cytotoxicity
- risk assessment
1. Introduction
2. Chemical Structure and Toxicity of PAs
PAs are mostly formed by a pyrrolizidine ring and an esterified organic acid, with the pyrrolizidine ring referred to as the necine and the acid part as the necic acid (Figure 1A). Based on the presence or absence of unsaturated double bonds at positions C-1 and C-2 of the necine structure, PAs are divided into saturated and unsaturated types. Saturated PAs with a saturated necine base, such as the platynecine (PLA) type, are known to be non-toxic. The unsaturated PAs are further divided into retronecine (RET), heliotridine (HEL), and otonecine (OTO) types (Figure 1B). OTO-type PAs include clivorine and senkirkine, while RET-type PAs include retrorsine and senecionine. HEL-type PAs include heliotrine and lasiocarpine (Figure 1C). Additionally, nitrogen atoms on the necine moiety can be oxidized to form N-oxides, which coexist with PAs in most plants [4]. However, otonecine-type PAs are not able to form a corresponding PA N-oxide due to the methylated nitrogen in the necine base core structure. Saturated PAs generally exhibit low or no toxicity, while 1,2-unsaturated PAs are of great concern due to their hepatotoxic, carcinogenic, and genotoxic properties [8].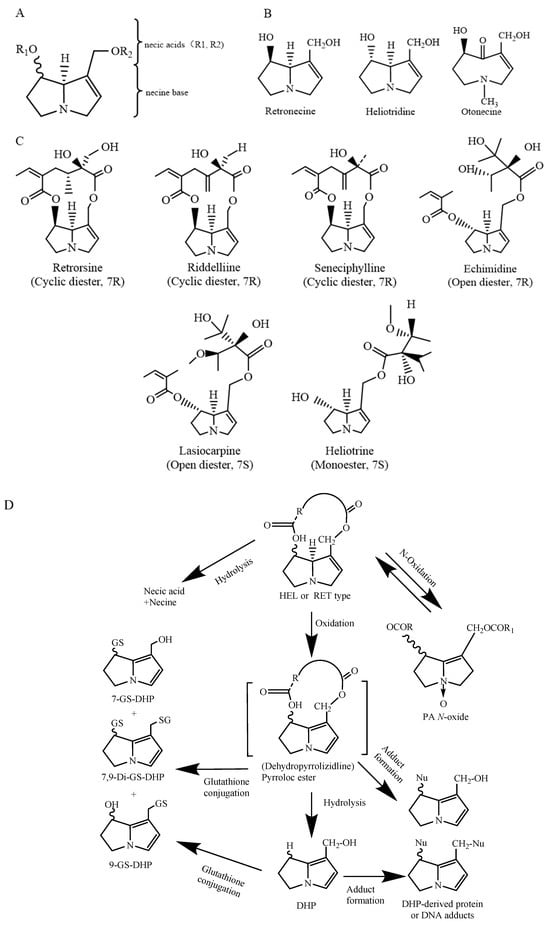
3. Toxic Effects of PAs
3.1. Acute Toxicity
Since the early 19th century, it has been observed that livestock that consume plants belonging to the genera Heliotropium, Senecio, or Crotalaria experience slow emaciation and weakness, and has autopsies have revealed hepatocyte necrosis [17][18][63,64]. Acute poisoning by PAs can significantly affect the liver, leading to acute veno-occlusive disease characterized by hepatomegaly, hemorrhage, ascites, and even death in severe cases [1]. It has been reported that a 6-month-old female infant was diagnosed with hepatic veno-occlusive disease (HVOD) after the ingestion of PAs at approximately 0.8 to 1.7 mg/kg body weight (b.w.) per day for 2 weeks [1]. Similarly, a 2-month-old male infant ingesting 3 mg/kg (b.w.) PAs per day died after approximately 4 days [19][65]. Based on epidemiological data, the EFSA Panel on Contaminants in the Food Chain estimated that the daily intake of PAs ranging from 1 to 3 mg/kg (b.w.) per day for 4 to 14 days can cause acute toxicity [8]. It was shown that 7R-configured macrocyclic diesters of PAs, including retrorsine, seneciphylline, and senecionine, constitute the most potent group causing acute toxicity [20][21][22][66,67,68]. The 7S-synthesized Pas heliotrine and lasiocarpine show acute toxic effects similar to macrocyclic diesters [20][66]. Compared with 7S-heliotrine, 7S-lasiocarpine, and 7R-configured macrocyclic diesters of PA, 7R-echimidine, 7S-heliotrine, 7R-indicine, and 7R-intermedine had lower acute toxic effects [23][69]. Furthermore, the acute toxicity of PANOs generally appears to be lower than that of their parent PAs [13].3.2. Cytotoxicity
The cytotoxicity induced by Pas is also structurally dependent. Li et al. (2013) evaluated the cytotoxicity of four PAs, namely, seneciphylline, senecionine, retrorsine, and riddelliine, on HepG2 cells using MTT and bromodeoxyuridine (BrdU) incorporation assays. MTT results showed that the IC20 value of senecionine was 0.66 mM, which was 2.4, 1.9, and 2.1 times greater than those of retrorsine, senecionine, and riddelliine, respectively. Moreover, the BrdU assay showed similar results [24][70]. Reuel A Field et al. (2015) assessed the effects of 11 PAs on cell morphology, mitochondrial function, and lactate dehydrogenase (LDH) activity in CRL-2118 chicken hepatoma cells. MTT and LDH assays revealed that lasiocarpine had the greatest cytotoxicity, followed by riddelliine, heliotrine, seneciphylline, and senecionine. The cytotoxic effects of these PAs are characterized by significant cell swelling and vacuoles. On the other hand, the cytotoxicities of riddelliine N-oxide, senecionine N-oxide, and heliotrine N-oxide were lower than those of monocrotaline, intermedine, and lycopsamine [25][71]. In general, the cytotoxicity caused by macrocyclic diesters (RET and HEL types) with cyclic and acyclic ester structures, such as lasiocarpine, seneciphylline, and riddelliine, was greater than that caused by monoesters (heliotrine, lycopsamine, and intermedine) and PANOs. Current studies on the cellular toxicity mechanisms of PAs have focused mainly on oxidative stress and apoptosis, with oxidative stress being the primary cause of cytotoxicity. Several studies revealed that DHPAs and DHP not only bind to DNA and proteins to form adducts but also bind to glutathione (GSH) to form adducts. When GSH is depleted and not supplied in a timely manner, it leads to oxidative stress and consequent cytotoxicity [26][72]. Previous studies have shown that exposure of rat hepatocytes to adonifoline, monocrotaline, and clivorine significantly reduces intracellular GSH levels, and the activities of glutathione peroxidase (GSH-Px), glutathione reductase (GR), and glutathione S-transferase (GST) also significantly decrease [26][27][72,73]. In addition, apoptosis is another crucial factor contributing to cytotoxicity. Several studies have demonstrated that PAs can cause hepatotoxicity by activating apoptosis [28][74]. Clivorine treatment of L-02 cells resulted in a significant increase in intracellular caspase-3 enzyme activity and increased expression levels of cleaved PARP and cleaved caspase-3 proteins [29][30][75,76]. Furthermore, the induction of apoptosis and subsequent cytotoxicity can also be attributed to the downregulation of the antiapoptotic factor Bcl-xl and the upregulation of Fas expression [29][31][75,77]. Table 1 summarizes the IC50 and IC20 values for cytotoxicity caused by different types of PAs.|
PAs |
Cell Line |
Exposure Dose (μM) |
Exposure Time (h) |
IC50/IC20 (μM) |
References |
|---|---|---|---|---|---|
|
Seneciphylline |
HepG2 |
62.5, 125, 250, 500, 1000 |
24 |
660 a |
|
|
Clivorine |
130 a |
||||
|
Retrorsine |
270 a |
||||
|
Platyphylline |
850 a |
||||
|
Senecionine |
340 a |
||||
|
Lasiocarpine |
CLR-2118 |
19, 38, 75, 300 |
24 |
14 b |
|
|
Senecionine |
mouse primary hepatocytes |
1, 3, 10, 30, 100 |
48 |
5.41 b |
|
|
Adonifoline |
49.91 b |
a IC20 b IC50; IC20 values refer to the calculated 20% inhibitory concentrations on cell viability; IC50 values refer to half-maximal inhibitory concentrations on cell viability.